Вам также может понравиться
- Increased COVID-19 Infections in Women With Polycystic Ovary Syndrome: A Population-Based StudyДокумент9 страницIncreased COVID-19 Infections in Women With Polycystic Ovary Syndrome: A Population-Based StudyDrFarah Emad AliОценок пока нет
- Kutz Et Al., 2021Документ10 страницKutz Et Al., 2021madelineОценок пока нет
- Brochure Basicsofalz LowДокумент28 страницBrochure Basicsofalz LowIbrahim HalilovicОценок пока нет
- Multiple Choice Questions on Androgen Production and EffectsДокумент4 страницыMultiple Choice Questions on Androgen Production and EffectsDrFarah Emad AliОценок пока нет
- Ovulation Test: 4 or MoreДокумент12 страницOvulation Test: 4 or MoreDrFarah Emad AliОценок пока нет
- Pancreas: Anatomy, Diseases and Health Implications: September 2012Документ53 страницыPancreas: Anatomy, Diseases and Health Implications: September 2012DrFarah Emad AliОценок пока нет
- Ovulation and Fertilisation and PlacentaДокумент28 страницOvulation and Fertilisation and PlacentaDrFarah Emad AliОценок пока нет
- Metabolism Open: Abu Saleh MD Moin, Thozhukat Sathyapalan, Stephen L. Atkin, Alexandra E. ButlerДокумент4 страницыMetabolism Open: Abu Saleh MD Moin, Thozhukat Sathyapalan, Stephen L. Atkin, Alexandra E. ButlerDrFarah Emad AliОценок пока нет
- Ovulation Test: 5 Important Points You Need To ReadДокумент2 страницыOvulation Test: 5 Important Points You Need To ReadDrFarah Emad AliОценок пока нет
- Introduction To Medical and Molecular BiologyДокумент95 страницIntroduction To Medical and Molecular BiologyJohnson KendekОценок пока нет
- Pancreas: Anatomy, Diseases and Health Implications: September 2012Документ53 страницыPancreas: Anatomy, Diseases and Health Implications: September 2012DrFarah Emad AliОценок пока нет
- Hepatic Circulation and Toxicology: Drug Metabolism Reviews February 1997Документ28 страницHepatic Circulation and Toxicology: Drug Metabolism Reviews February 1997DrFarah Emad AliОценок пока нет
- Physiology HypothalamusДокумент6 страницPhysiology HypothalamusDrFarah Emad AliОценок пока нет
- Technologies For Cervical Cancer Detection, Diagnosis, Monitoring and Treatment in Low-Resource SettingsДокумент42 страницыTechnologies For Cervical Cancer Detection, Diagnosis, Monitoring and Treatment in Low-Resource SettingsDrFarah Emad AliОценок пока нет
- Brochure Basicsofalz LowДокумент28 страницBrochure Basicsofalz LowIbrahim HalilovicОценок пока нет
- ObsGyn OSCE NotesДокумент52 страницыObsGyn OSCE Notesmysorewala100% (11)
- Cardiac Function Tests: A Body and Disease WorkshopДокумент32 страницыCardiac Function Tests: A Body and Disease WorkshopDrFarah Emad AliОценок пока нет
- Doses of Misoprostol Use Updated March 2018 PDFДокумент2 страницыDoses of Misoprostol Use Updated March 2018 PDFDrFarah Emad AliОценок пока нет
- Cognitive Assessment Tip SheetДокумент1 страницаCognitive Assessment Tip SheetDrFarah Emad AliОценок пока нет
- Renal Function TestsДокумент19 страницRenal Function TestsDrFarah Emad AliОценок пока нет
- Thyroid Gland and Its Rule in Human BodyДокумент9 страницThyroid Gland and Its Rule in Human BodyDrFarah Emad AliОценок пока нет
- Risk Assessment For Venous ThromboembolismДокумент1 страницаRisk Assessment For Venous ThromboembolismDrFarah Emad AliОценок пока нет
- Mol Path-Clinician Collected Vag SwabДокумент1 страницаMol Path-Clinician Collected Vag SwabDevira ParamithaОценок пока нет
- Chapter-25 Kidney Function Tests: January 2017Документ14 страницChapter-25 Kidney Function Tests: January 2017DrFarah Emad AliОценок пока нет
- Ovulation and Fertilisation and PlacentaДокумент28 страницOvulation and Fertilisation and PlacentaDrFarah Emad AliОценок пока нет
- COVID-19 Infection & Woman HealthДокумент17 страницCOVID-19 Infection & Woman HealthDrFarah Emad AliОценок пока нет
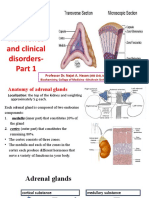
- Adrenal Gland: Structure, Hormones and Clinical Disorders-: Professor Dr. Najat A. HasanДокумент18 страницAdrenal Gland: Structure, Hormones and Clinical Disorders-: Professor Dr. Najat A. HasanDrFarah Emad Ali100% (1)
- Depression and Multiple Sclerosis PDFДокумент9 страницDepression and Multiple Sclerosis PDFDrFarah Emad AliОценок пока нет
- Anti D Ig Prophylactic Use in Obstetrics Iraqi Protocol 2016Документ5 страницAnti D Ig Prophylactic Use in Obstetrics Iraqi Protocol 2016DrFarah Emad AliОценок пока нет
- Record of Achievement in OpenWHO TB CourseДокумент1 страницаRecord of Achievement in OpenWHO TB CourseDrFarah Emad AliОценок пока нет
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (119)
- Ashes Reborn - WikiДокумент27 страницAshes Reborn - WikiIvan StancheffОценок пока нет
- Shadowdark RPG - Welcome Pack - BWДокумент17 страницShadowdark RPG - Welcome Pack - BWVal89% (9)
- English Grammar Cloud 8Документ24 страницыEnglish Grammar Cloud 8Aismita BarОценок пока нет
- Issue 235 Organic Consumers AssociationДокумент1 страницаIssue 235 Organic Consumers AssociationFriends of Organic Consumers AssociationОценок пока нет
- Ujian Masuk Program Pascasarjana: Jangan Dibuka Dulu. Tunggu PetunjukДокумент21 страницаUjian Masuk Program Pascasarjana: Jangan Dibuka Dulu. Tunggu PetunjukLalu SuhaimiОценок пока нет
- Don't Say Forever Victoria Falls SequenceДокумент9 страницDon't Say Forever Victoria Falls SequenceMaha GolestanehОценок пока нет
- Present Tense VerbДокумент15 страницPresent Tense VerbMai Mai BallanoОценок пока нет
- Eucoelomates Lab ReportДокумент13 страницEucoelomates Lab Reportsatvindar singhОценок пока нет
- Mga Uri NG ManokДокумент31 страницаMga Uri NG ManokMa LeslynneОценок пока нет
- Plasma Proteins by U.Sivakumar: 1 PhysiologyДокумент23 страницыPlasma Proteins by U.Sivakumar: 1 PhysiologyAkash JaatОценок пока нет
- Colonic LesionsДокумент89 страницColonic Lesionsjagadeesan_ushaОценок пока нет
- OdomДокумент31 страницаOdomRangothri Sreenivasa SubramanyamОценок пока нет
- Fables Grade 2Документ4 страницыFables Grade 2angeliОценок пока нет
- Laying vs. Lying (Lay vs. Lie) - Learn It Easily - GrammarlyДокумент4 страницыLaying vs. Lying (Lay vs. Lie) - Learn It Easily - GrammarlyDavid Puente100% (1)
- 2023 Pink Schedule FinalДокумент20 страниц2023 Pink Schedule Finalapi-84174556Оценок пока нет
- Animal Farm / Chapter 8 Reading Organizer Sample AnswersДокумент7 страницAnimal Farm / Chapter 8 Reading Organizer Sample AnswersJacques SnicketОценок пока нет
- Paed HistДокумент123 страницыPaed HistG VenkateshОценок пока нет
- Kiara Maestre 2020 05 12 1420 PDFДокумент2 страницыKiara Maestre 2020 05 12 1420 PDFmarionochesОценок пока нет
- Book - Feed Me BillyДокумент10 страницBook - Feed Me BillyАсель РымхановаОценок пока нет
- CH 12 Word ListДокумент3 страницыCH 12 Word ListtigertiaОценок пока нет
- Cure - Family Health MatterzДокумент14 страницCure - Family Health MatterzGeorge AniborОценок пока нет
- Lounging Leafer Giraffe DollДокумент33 страницыLounging Leafer Giraffe DollDana Richards100% (1)
- Obada - 2010 - Systematic Atribution of The Most Ancient ElephantidaeДокумент29 страницObada - 2010 - Systematic Atribution of The Most Ancient ElephantidaeTheodor Obada100% (1)
- The Complex SaraДокумент10 страницThe Complex SaraFlame10100% (3)
- Phrases and SentencesДокумент39 страницPhrases and SentencesGhie Yambao SarmientoОценок пока нет
- Indian import duty rates for live animals, meat, fish and aquatic productsДокумент98 страницIndian import duty rates for live animals, meat, fish and aquatic productsGopal UОценок пока нет
- Script Jungle CruiseДокумент24 страницыScript Jungle CruiseronnyОценок пока нет
- Understanding Poultry Game BirdsДокумент33 страницыUnderstanding Poultry Game BirdsThoyyibОценок пока нет
- Subject: Telugu Tejam FactsДокумент5 страницSubject: Telugu Tejam FactsAum RaoОценок пока нет
- Animal Trafficking Lesson PlanДокумент4 страницыAnimal Trafficking Lesson Plankurucz barbaraОценок пока нет