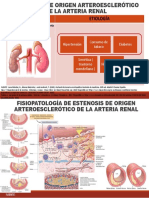
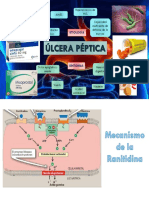
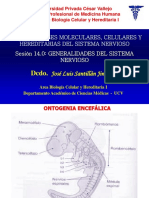
Вам также может понравиться
- PPTДокумент2 страницыPPTVictor Pier Chuquipoma RojasОценок пока нет
- Guía de Prácticas #05 PDFДокумент5 страницGuía de Prácticas #05 PDFVictor Pier Chuquipoma RojasОценок пока нет
- Biosíntesis de Estrogenos BioloДокумент2 страницыBiosíntesis de Estrogenos BioloVictor Pier Chuquipoma RojasОценок пока нет
- 002 MAIS - Principios Integradores 2017-2Документ27 страниц002 MAIS - Principios Integradores 2017-2Victor Pier Chuquipoma RojasОценок пока нет
- Semana 1Документ2 страницыSemana 1Victor Pier Chuquipoma RojasОценок пока нет
- Churcampa AvancesДокумент17 страницChurcampa AvancesVictor Pier Chuquipoma RojasОценок пока нет
- Semana 1Документ2 страницыSemana 1Victor Pier Chuquipoma RojasОценок пока нет
- Funda ExpoДокумент4 страницыFunda ExpoVictor Pier Chuquipoma RojasОценок пока нет
- Insuficiencia Respiratoria: Definición EtiologíaДокумент4 страницыInsuficiencia Respiratoria: Definición EtiologíaVictor Pier Chuquipoma RojasОценок пока нет
- Caso ClicnicoДокумент18 страницCaso ClicnicoVictor Pier Chuquipoma RojasОценок пока нет
- Silabo - Salud PúblicaДокумент11 страницSilabo - Salud PúblicaVictor Pier Chuquipoma RojasОценок пока нет
- Glándula Tiroides y ParatiroidesДокумент5 страницGlándula Tiroides y ParatiroidesVictor Pier Chuquipoma RojasОценок пока нет
- TiposPirámidesPoblaciónДокумент8 страницTiposPirámidesPoblaciónpablovillar76Оценок пока нет
- Analisis Del Centro de Salud Victor LarcoДокумент60 страницAnalisis Del Centro de Salud Victor LarcoVictor Pier Chuquipoma Rojas50% (2)
- Descentralizacion y Gestion TerritorialДокумент14 страницDescentralizacion y Gestion TerritorialVictor Pier Chuquipoma RojasОценок пока нет
- Gu A Taller Instrumentos de FamiliaДокумент13 страницGu A Taller Instrumentos de FamiliaFurboo Toretto ValdezОценок пока нет
- Diapositivas Redaccion UniversitariasДокумент17 страницDiapositivas Redaccion UniversitariasVictor Pier Chuquipoma RojasОценок пока нет
- RM 204 2015 MINSA Ficha Familiar.Документ7 страницRM 204 2015 MINSA Ficha Familiar.susanОценок пока нет
- FamiliaДокумент6 страницFamiliaCaro De Lima MalladaОценок пока нет
- 5336material Profesores UD 1 - Funciones y RolesДокумент23 страницы5336material Profesores UD 1 - Funciones y RolesVictor Pier Chuquipoma RojasОценок пока нет
- Sistema Digestivo Clase 1 Dr. BarónДокумент38 страницSistema Digestivo Clase 1 Dr. BarónVictor Pier Chuquipoma RojasОценок пока нет
- SOCIODEMOGRAFIAДокумент8 страницSOCIODEMOGRAFIAitmi4Оценок пока нет
- Mecanismo Meiosis Clase 8 Dr. BarónДокумент41 страницаMecanismo Meiosis Clase 8 Dr. BarónVictor Pier Chuquipoma RojasОценок пока нет
- Semana 1Документ2 страницыSemana 1Victor Pier Chuquipoma RojasОценок пока нет
- Sistema Urinario Clase 6 Dr. BarónДокумент34 страницыSistema Urinario Clase 6 Dr. BarónVictor Pier Chuquipoma RojasОценок пока нет
- Meiosis I IIДокумент22 страницыMeiosis I IIVictor Pier Chuquipoma RojasОценок пока нет
- Sistema Digestivo Clase 3 Dr. BarónДокумент44 страницыSistema Digestivo Clase 3 Dr. BarónVictor Pier Chuquipoma RojasОценок пока нет
- Sistema Urinario Clase 5 Dr. BarónДокумент37 страницSistema Urinario Clase 5 Dr. BarónVictor Pier Chuquipoma RojasОценок пока нет
- Duplicción Transcripción Código Genetico TraducciónДокумент19 страницDuplicción Transcripción Código Genetico TraducciónVictor Pier Chuquipoma RojasОценок пока нет
- Generalidades Sistema NerviosoДокумент34 страницыGeneralidades Sistema NerviosoVictor Pier Chuquipoma RojasОценок пока нет
- Monografia D Oncogenes y ProtooncogenesДокумент11 страницMonografia D Oncogenes y ProtooncogenesCesar Huallpa0% (1)
- PlanTrabajoBIOQUÍMICA IIДокумент8 страницPlanTrabajoBIOQUÍMICA IIdoyneldavidОценок пока нет
- TP 2 Mitosis y MeiosisДокумент7 страницTP 2 Mitosis y MeiosisDen DiazОценок пока нет
- Los ParpsДокумент10 страницLos ParpsAlejandro ReyesОценок пока нет
- Tabla de Cofactores InorganicosДокумент3 страницыTabla de Cofactores Inorganicosyessenia apaza mamaniОценок пока нет
- Capítulo 5. PseudogenesДокумент9 страницCapítulo 5. PseudogenesJuan Manuel MejíaОценок пока нет
- Ejercicios Cinética Enzimática Bioquímica U2019Документ1 страницаEjercicios Cinética Enzimática Bioquímica U2019Jenly RamírezОценок пока нет
- 2 Nucleoide BacterianoДокумент13 страниц2 Nucleoide Bacterianoeduardo enrique thorrens romeroОценок пока нет
- MeiosisДокумент3 страницыMeiosisMelissa Berrocal BarbozaОценок пока нет
- Introducción Al MetabolismoДокумент26 страницIntroducción Al MetabolismoAlessandra CotrinaОценок пока нет
- Sistemas energéticos deporteДокумент83 страницыSistemas energéticos deportedianaОценок пока нет
- Respiración Celular 2023Документ15 страницRespiración Celular 2023alansegundo400Оценок пока нет
- Embriologia y Genetica - Grupo 4 - Factores de Señalización ParacrinaДокумент9 страницEmbriologia y Genetica - Grupo 4 - Factores de Señalización ParacrinaKathlen SanchezОценок пока нет
- SirtuinasДокумент57 страницSirtuinasΜαρία ΛόπεζОценок пока нет
- Cómo obtienen energía las células a través del metabolismo celularДокумент2 страницыCómo obtienen energía las células a través del metabolismo celularRodrigo Calisaya PortillaОценок пока нет
- Código GenéticoДокумент5 страницCódigo GenéticoMiguel Angel Rodas HerreraОценок пока нет
- Biologia 06 Genetica y Metabolismo CelularДокумент3 страницыBiologia 06 Genetica y Metabolismo Celularrami pinanОценок пока нет
- Enzimas aceleran reacciones metabólicasДокумент3 страницыEnzimas aceleran reacciones metabólicasPelukas Riveros ArceОценок пока нет
- Taller Final SBДокумент3 страницыTaller Final SBbetincarlos3010Оценок пока нет
- Diapositivas de Fotosíntesis Fase LuminosaДокумент16 страницDiapositivas de Fotosíntesis Fase LuminosaALI RACHEL SALGADO GAFFARE 3BVLОценок пока нет
- Ciclo de KrebsДокумент4 страницыCiclo de KrebsCesar Eugenio Sanhueza ValdebenitoОценок пока нет
- Biología celular y molecular: Ciclo celular, ácidos nucleicos y muerte celularДокумент18 страницBiología celular y molecular: Ciclo celular, ácidos nucleicos y muerte celularSandra Paola Fonseca Acosta100% (1)
- Cold Spring Harb Perspect Biol-2021-Chandel-a040535.en - EsДокумент12 страницCold Spring Harb Perspect Biol-2021-Chandel-a040535.en - EsValeria Flores HiriartОценок пока нет
- Seminario 04 - DesarrolladoДокумент5 страницSeminario 04 - DesarrolladoAísha VkОценок пока нет
- Ciclo celular: Fases, Regulación y ControlДокумент8 страницCiclo celular: Fases, Regulación y ControlPablo Jorquera GarcésОценок пока нет
- Podcast KrebsДокумент2 страницыPodcast Krebseliezer montesОценок пока нет
- Ciclo de KrebsДокумент6 страницCiclo de KrebsAguila PaulОценок пока нет
- PAUTA Taller Ciclo Celular BIOL 034 Reforzamiento ProfesoresДокумент7 страницPAUTA Taller Ciclo Celular BIOL 034 Reforzamiento Profesoresjaviera rivera ramirezОценок пока нет
- Acidos NucleicosДокумент2 страницыAcidos NucleicosAMIRA DESSYREE DUARTE NUÑEZОценок пока нет
- Copia de Lectura - AutophagyДокумент6 страницCopia de Lectura - AutophagyzzzzzzzzОценок пока нет