Вам также может понравиться
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Can - Can't ActivitiesДокумент2 страницыCan - Can't ActivitiesRalucaTeodorescuОценок пока нет
- Shapes LessonДокумент2 страницыShapes LessonRalucaTeodorescuОценок пока нет
- Lesson 5Документ2 страницыLesson 5RalucaTeodorescuОценок пока нет
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Lesson 3Документ3 страницыLesson 3RalucaTeodorescuОценок пока нет
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- House ObjectsДокумент3 страницыHouse ObjectsRalucaTeodorescuОценок пока нет
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
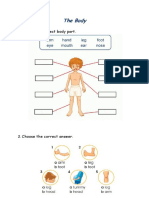
- The Body Have-Has GotДокумент3 страницыThe Body Have-Has GotRalucaTeodorescuОценок пока нет
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- The Portrayal of Minority Women in The Color Purple and BelovedДокумент24 страницыThe Portrayal of Minority Women in The Color Purple and BelovedRalucaTeodorescuОценок пока нет
- Elementary Unit 2 Video Podcast Script: PHOTOCOPIABLE © 2011 Pearson LongmanДокумент2 страницыElementary Unit 2 Video Podcast Script: PHOTOCOPIABLE © 2011 Pearson LongmanRalucaTeodorescuОценок пока нет
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Elementary Unit 8 Video Podcast Script: Trek, I Think, Star Trek 5, I Think It WasДокумент2 страницыElementary Unit 8 Video Podcast Script: Trek, I Think, Star Trek 5, I Think It WasRalucaTeodorescuОценок пока нет
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- Elementary Unit 1 Video Podcast ScriptДокумент3 страницыElementary Unit 1 Video Podcast ScriptRalucaTeodorescuОценок пока нет
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Elementary Unit 3 Video Podcast ScriptДокумент2 страницыElementary Unit 3 Video Podcast ScriptRalucaTeodorescuОценок пока нет
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- The Washington Post 1Документ14 страницThe Washington Post 1RalucaTeodorescuОценок пока нет
- Elementary Unit 4 Video Podcast Script: PHOTOCOPIABLE © 2011 Pearson LongmanДокумент2 страницыElementary Unit 4 Video Podcast Script: PHOTOCOPIABLE © 2011 Pearson LongmanRalucaTeodorescuОценок пока нет
- DEFENSIVE MECHANISMS OF PLANTS BASED ON SECONDARY METABOLITES - Alexan DianaДокумент8 страницDEFENSIVE MECHANISMS OF PLANTS BASED ON SECONDARY METABOLITES - Alexan DianaRalucaTeodorescuОценок пока нет
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Enfranchisement EuДокумент6 страницEnfranchisement EuRalucaTeodorescuОценок пока нет
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- AntiFederalist Centinel 1 PDFДокумент8 страницAntiFederalist Centinel 1 PDFRalucaTeodorescuОценок пока нет
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- Essay Assignment IДокумент2 страницыEssay Assignment IRalucaTeodorescuОценок пока нет
- AntiFederalist Centinel 1Документ8 страницAntiFederalist Centinel 1RalucaTeodorescuОценок пока нет
- Router SettingsДокумент1 страницаRouter SettingsRalucaTeodorescuОценок пока нет
- Essay Final DraftДокумент10 страницEssay Final DraftRalucaTeodorescuОценок пока нет
- Early Essays On Marriage and DivorceДокумент2 страницыEarly Essays On Marriage and DivorceRalucaTeodorescuОценок пока нет
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Harriet Taylor MillДокумент11 страницHarriet Taylor MillRalucaTeodorescuОценок пока нет
- Essay PercecДокумент3 страницыEssay PercecRalucaTeodorescuОценок пока нет
- Essay DraftДокумент10 страницEssay DraftRalucaTeodorescuОценок пока нет
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- Basic 1: Personal PronounsДокумент3 страницыBasic 1: Personal PronounsRalucaTeodorescuОценок пока нет
- Teodorescu Frankfurt SchoolДокумент13 страницTeodorescu Frankfurt SchoolRalucaTeodorescuОценок пока нет
- Raluca Teodorescu, First DraftДокумент7 страницRaluca Teodorescu, First DraftRalucaTeodorescuОценок пока нет
- Enfranchisement EuДокумент6 страницEnfranchisement EuRalucaTeodorescuОценок пока нет
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (120)
- Alternative Words For Effective WritingДокумент5 страницAlternative Words For Effective Writingmeenadeepak007Оценок пока нет
- Fracture Characterization of Threaded Rebar and Coupler: February 2016Документ5 страницFracture Characterization of Threaded Rebar and Coupler: February 2016Hotel Green DotОценок пока нет
- Quality AssuranceДокумент8 страницQuality AssuranceEmmanuelОценок пока нет
- 30.sun Lipstick 0.270.01 C100 27112015Документ1 страница30.sun Lipstick 0.270.01 C100 27112015azarine wahanaОценок пока нет
- TS 15280Документ1 страницаTS 15280GiveawayОценок пока нет
- Steels ExplainedДокумент237 страницSteels Explainedradou99100% (1)
- As1 Cee317b 2022Документ4 страницыAs1 Cee317b 2022Pheletso Andrias MoloantoaОценок пока нет
- Biomass FinalДокумент12 страницBiomass FinalAbu ikhtiarОценок пока нет
- Science and Engineering of Materials 7th Edition Askeland Solutions ManualДокумент29 страницScience and Engineering of Materials 7th Edition Askeland Solutions Manualcrapevioloush1o97% (32)
- Solitary Waves and Supersonic Reaction Front in Metastable SolidsДокумент30 страницSolitary Waves and Supersonic Reaction Front in Metastable Solidstestonly261Оценок пока нет
- Phil 110 Chemical Reaction EssayДокумент5 страницPhil 110 Chemical Reaction Essayapi-253261499Оценок пока нет
- Comparison of The Use of Rutile and Cellulosic Electrodes - TWIДокумент11 страницComparison of The Use of Rutile and Cellulosic Electrodes - TWISumantaОценок пока нет
- CATALOG Uniweld-Products TYPE17-2Документ13 страницCATALOG Uniweld-Products TYPE17-2Máy cầm tay Hoàng TrungОценок пока нет
- 3.05 Ionic Bonding and Ionic CompoundsДокумент3 страницы3.05 Ionic Bonding and Ionic CompoundsIBRAHIM ABOU EL NAAJОценок пока нет
- Guide To BondingДокумент46 страницGuide To Bondingbotan_mihaiОценок пока нет
- Le Ngoc Lieu - Chapter 1 - DehydrationДокумент117 страницLe Ngoc Lieu - Chapter 1 - DehydrationThuỳ TrangОценок пока нет
- Materials of Construction and Selection: Faculty of Chemical Engineering Universiti Teknologi MaraДокумент80 страницMaterials of Construction and Selection: Faculty of Chemical Engineering Universiti Teknologi MaraAisyah Addia AzizanОценок пока нет
- Assignment 1 - CarbohydratesДокумент4 страницыAssignment 1 - CarbohydratesJay Ann BernalesОценок пока нет
- Unit V BIO-CHEMICAL MEASUREMENTДокумент8 страницUnit V BIO-CHEMICAL MEASUREMENTAleeshaОценок пока нет
- Catalogue CSM PDFДокумент4 страницыCatalogue CSM PDFFran IgledominguezОценок пока нет
- BL4S Results 2021Документ2 страницыBL4S Results 2021Sadika AkhterОценок пока нет
- Caustic Management and Reuse in The Beverage Bottling IndustryДокумент52 страницыCaustic Management and Reuse in The Beverage Bottling IndustrydingobkОценок пока нет
- Syntheses Vanillyl AlcoholsДокумент48 страницSyntheses Vanillyl AlcoholsIlmuncMakesuill100% (1)
- Els Week 1Документ13 страницEls Week 1Paolin Pearl Ortiola RapisuraОценок пока нет
- FOB Megaflow AW Hydraulic Oil TDSW 814637aДокумент3 страницыFOB Megaflow AW Hydraulic Oil TDSW 814637aElwin arifinОценок пока нет
- Pattern Shop InstructionsДокумент19 страницPattern Shop InstructionsSaif UllahОценок пока нет
- Hectatone Universal ProductBulletinДокумент1 страницаHectatone Universal ProductBulletinFederico ArqueroОценок пока нет
- Protein-Synthesis WorksheetДокумент5 страницProtein-Synthesis WorksheetAme RealОценок пока нет
- Research Papers Structure of The Novel Ternary Hydrides Li TT D (TT Si and Ge)Документ6 страницResearch Papers Structure of The Novel Ternary Hydrides Li TT D (TT Si and Ge)Fadjar MulyaОценок пока нет
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouОт EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouРейтинг: 4.5 из 5 звезд4.5/5 (62)
- Why We Die: The New Science of Aging and the Quest for ImmortalityОт EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityРейтинг: 4 из 5 звезд4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisОт EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisРейтинг: 3.5 из 5 звезд3.5/5 (2)
- Tales from Both Sides of the Brain: A Life in NeuroscienceОт EverandTales from Both Sides of the Brain: A Life in NeuroscienceРейтинг: 3 из 5 звезд3/5 (18)
- Gut: the new and revised Sunday Times bestsellerОт EverandGut: the new and revised Sunday Times bestsellerРейтинг: 4 из 5 звезд4/5 (392)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedОт EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedРейтинг: 4 из 5 звезд4/5 (11)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceОт EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceРейтинг: 4.5 из 5 звезд4.5/5 (516)