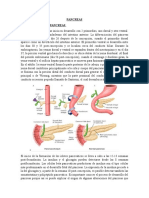
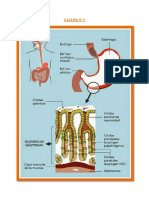
Вам также может понравиться
- LAPAROTOMIAДокумент155 страницLAPAROTOMIAJohnAnthonySeguraAltamirano100% (1)
- Anatomia PancreasДокумент30 страницAnatomia PancreasDianaAvendañoHuaychaniОценок пока нет
- El Hígado Anatomía y FisiologíaДокумент13 страницEl Hígado Anatomía y FisiologíaRoberto ChacónОценок пока нет
- El P Ncreas Es Una GL Ndula Mixta Endocrina y ExoДокумент2 страницыEl P Ncreas Es Una GL Ndula Mixta Endocrina y Exojose benitezОценок пока нет
- PancreatitisДокумент48 страницPancreatitisRicardo Avila Saavedra100% (1)
- Taller 2. Biologia Celular y MolecularДокумент7 страницTaller 2. Biologia Celular y MolecularMARIA LUCIA PIESCHACÓN CHUZCANOОценок пока нет
- Universidad Nacional Del Santa Facultad de Ciencias de La Salud Escuela Profesional de Medicina Humana Fisiología Humana IiДокумент4 страницыUniversidad Nacional Del Santa Facultad de Ciencias de La Salud Escuela Profesional de Medicina Humana Fisiología Humana IiYanina Vasquez NuñezОценок пока нет
- Hormonas PancreaticasДокумент18 страницHormonas PancreaticasTenaz100% (2)
- Orgánulos Celulares 2º BachДокумент4 страницыOrgánulos Celulares 2º BachMónica Fernández SantosОценок пока нет
- Guía Diversidad Células HumanasДокумент17 страницGuía Diversidad Células HumanasMartha VegaОценок пока нет
- Corteza Suprarrenal y HM PancreáticasДокумент39 страницCorteza Suprarrenal y HM Pancreáticasclases.trioОценок пока нет
- 45 46 Merged RemovedДокумент19 страниц45 46 Merged RemovedWendy BojorquezОценок пока нет
- Guia de Estudio - CompartimientosДокумент36 страницGuia de Estudio - Compartimientosnatalia.perez.camigliaОценок пока нет
- Ejercicio de BiomoleculasДокумент30 страницEjercicio de BiomoleculasOregel Castrejón ValeriaОценок пока нет
- Metabolismo de Las Lipoproteínas PDFДокумент9 страницMetabolismo de Las Lipoproteínas PDFWendell CallenderОценок пока нет
- Cap.10 Orgánulos Que Participan en La Sintesis y DegradacionДокумент21 страницаCap.10 Orgánulos Que Participan en La Sintesis y DegradacionJuan Carlos Ledo GarciaОценок пока нет
- HEPATOCITOSДокумент4 страницыHEPATOCITOSDaniela TenorioОценок пока нет
- Fisiología Del HígadoДокумент27 страницFisiología Del HígadoGerardoОценок пока нет
- ANEXOSДокумент3 страницыANEXOSGABRIELA ADRIANA LEON MERINOОценок пока нет
- Taller 3 An 1Документ9 страницTaller 3 An 1angienataliatenorio96Оценок пока нет
- Degradacion de Proteinas ExposicionДокумент5 страницDegradacion de Proteinas ExposicionMara Paola DurangoОценок пока нет
- 2da Charla CirugiaДокумент4 страницы2da Charla CirugiaEstela QuirosОценок пока нет
- Capítulo 1 LA CELULA COMO UNIDAD DE SALUD Y ENFERMEDADДокумент6 страницCapítulo 1 LA CELULA COMO UNIDAD DE SALUD Y ENFERMEDADMaria Camila González100% (1)
- Pancreas ExocrinoДокумент8 страницPancreas ExocrinoDiego GervacioОценок пока нет
- Fundamentos de La Fisiología Celular IДокумент3 страницыFundamentos de La Fisiología Celular IPaola Vianey Gonzalez NavarroОценок пока нет
- Páncreas y Vesícula BiliarДокумент6 страницPáncreas y Vesícula BiliarOsmar CorveraОценок пока нет
- UP2 - CyD 2020Документ29 страницUP2 - CyD 2020Benet Sant'anaОценок пока нет
- Células HepáticasДокумент3 страницыCélulas HepáticasletileoОценок пока нет
- Glanulas y Secrecion - ViДокумент20 страницGlanulas y Secrecion - VihottizdssОценок пока нет
- Celula EucariotaДокумент12 страницCelula Eucariotamarta SanchezОценок пока нет
- Endocrinologia 2018 IIДокумент34 страницыEndocrinologia 2018 IIAlisson SanchezОценок пока нет
- Resumen Citología, Estructura y Función CelularДокумент2 страницыResumen Citología, Estructura y Función CelularKatherine Beraún60% (5)
- PDF UP2 - NuevoДокумент24 страницыPDF UP2 - NuevonalaОценок пока нет
- Diabetes MellitusДокумент51 страницаDiabetes Mellitusana100% (1)
- PANCREATITISДокумент10 страницPANCREATITISMichelle AvalosОценок пока нет
- Resumen de Gyton Cap 1Документ57 страницResumen de Gyton Cap 1LeyDi CamposanoОценок пока нет
- Páncreas Exposición HistologiaДокумент5 страницPáncreas Exposición HistologiaceliaacostazepОценок пока нет
- PEROXISOMASДокумент32 страницыPEROXISOMASYaser Axel Alzamora ManosalvaОценок пока нет
- Retículo EndoplamásticoДокумент8 страницRetículo EndoplamásticoJorge Mendoza DiosesОценок пока нет
- El Sistema de Endomembranas - pptpara2010.Pptcap.VДокумент138 страницEl Sistema de Endomembranas - pptpara2010.Pptcap.VlalitotОценок пока нет
- Biología Tema 8. La Célula Eucariota Orgánulos MembranososДокумент7 страницBiología Tema 8. La Célula Eucariota Orgánulos MembranososAnaОценок пока нет
- Informe Re GolgiДокумент7 страницInforme Re GolgienriqueОценок пока нет
- Tarea Evalutiva PDFДокумент2 страницыTarea Evalutiva PDFYiliet Rodriguez QuevedoОценок пока нет
- Super Resumen PancreasДокумент12 страницSuper Resumen PancreasJheferson Alejandro Martinez Castillo100% (1)
- Unidad 2Документ41 страницаUnidad 2Matias VillagranОценок пока нет
- Guia de Estudio COMPARTIMIENTOS INTRACELULARESДокумент47 страницGuia de Estudio COMPARTIMIENTOS INTRACELULARESpimenteltamara2Оценок пока нет
- Taller N5Документ12 страницTaller N5Grecia AlvarezОценок пока нет
- Cecm00055 421 440Документ20 страницCecm00055 421 440Anna PeiruОценок пока нет
- Estructura y Función de Los Organelos CelularesДокумент4 страницыEstructura y Función de Los Organelos CelularesJosé Daniel Canchila0% (1)
- Organelos Según La Función Que Cumplan: SíntesisДокумент56 страницOrganelos Según La Función Que Cumplan: SíntesisCARLOS VERGARA COQUEОценок пока нет
- Cuestionario de BiologíaДокумент5 страницCuestionario de Biologíalolcitomobile5Оценок пока нет
- Resumen Páncreas Finn GeneserДокумент3 страницыResumen Páncreas Finn GeneserLaura FMОценок пока нет
- Sistema Neuroendocrino Difuso1Документ10 страницSistema Neuroendocrino Difuso1Vanesa De Los SantosОценок пока нет
- Protocolo Resumen de RossДокумент188 страницProtocolo Resumen de RossMi GoBer100% (1)
- HepatocitosДокумент5 страницHepatocitosMichael Macdonald50% (4)
- Resumen de RossДокумент188 страницResumen de RossRaul Melendez Navarro100% (13)
- Rejuvenecer Con El Plasma Sanguíneo De Los JóvenesОт EverandRejuvenecer Con El Plasma Sanguíneo De Los JóvenesРейтинг: 5 из 5 звезд5/5 (1)
- Metabolismo del sistema digestivo, del hígado, de la vesícula y de las vías biliares: En condiciones de salud y en las enfermedadesОт EverandMetabolismo del sistema digestivo, del hígado, de la vesícula y de las vías biliares: En condiciones de salud y en las enfermedadesОценок пока нет
- Higiene y Ambiente SaludableДокумент14 страницHigiene y Ambiente SaludableMaria Isabel Llamoga MiñanoОценок пока нет
- Toxoplasmosis CerebralДокумент5 страницToxoplasmosis CerebralMaria Isabel Llamoga MiñanoОценок пока нет
- DiverticulosisДокумент9 страницDiverticulosisMaria Isabel Llamoga MiñanoОценок пока нет
- Pseudomonas SP y AeruginosasДокумент13 страницPseudomonas SP y AeruginosasMaria Isabel Llamoga MiñanoОценок пока нет
- Staphylococcus SaprophyticusДокумент6 страницStaphylococcus SaprophyticusMaria Isabel Llamoga MiñanoОценок пока нет
- 10-19-2019 101324 Am UROCultivo y AntibiogramaДокумент43 страницы10-19-2019 101324 Am UROCultivo y AntibiogramaMaria Isabel Llamoga MiñanoОценок пока нет
- MicrobologíaДокумент11 страницMicrobologíaMaria Isabel Llamoga MiñanoОценок пока нет
- Staphylococcus SaprophyticusДокумент6 страницStaphylococcus SaprophyticusMaria Isabel Llamoga MiñanoОценок пока нет
- Staphylococcus SaprophyticusДокумент6 страницStaphylococcus SaprophyticusMaria Isabel Llamoga MiñanoОценок пока нет
- Laboratorio (Dislipidemia)Документ5 страницLaboratorio (Dislipidemia)Maria Isabel Llamoga MiñanoОценок пока нет
- KlebsiellaДокумент3 страницыKlebsiellaMaria Isabel Llamoga MiñanoОценок пока нет
- Estreptococos y Enterococos VD PDFДокумент89 страницEstreptococos y Enterococos VD PDFMaria Isabel Llamoga MiñanoОценок пока нет
- Caso ClínicoДокумент107 страницCaso ClínicoMaria Isabel Llamoga MiñanoОценок пока нет
- Staphylococcus SaprophyticusДокумент4 страницыStaphylococcus SaprophyticusMaria Isabel Llamoga Miñano0% (1)
- 07 - Relación Entre Ética y CiudadaníaДокумент17 страниц07 - Relación Entre Ética y CiudadaníaMaria Isabel Llamoga MiñanoОценок пока нет
- Informe 12 Diuréticos en CobayosДокумент17 страницInforme 12 Diuréticos en CobayosMaria Isabel Llamoga MiñanoОценок пока нет
- Limpieza HepáticaДокумент3 страницыLimpieza HepáticaposanterОценок пока нет
- 3 PancreasДокумент120 страниц3 PancreasArnoldTitoОценок пока нет
- Feline Hepatic Lipidosis2Документ20 страницFeline Hepatic Lipidosis2Nicole FuentesОценок пока нет
- Semiologiadeabdomen Parte 1Документ73 страницыSemiologiadeabdomen Parte 1Jefferson IsraelОценок пока нет
- CG01 - Vesícula y Vías BiliaresДокумент13 страницCG01 - Vesícula y Vías BiliaresGabriel Rojas CastroОценок пока нет
- Higado, Pancreas y Vesicula BiliarДокумент69 страницHigado, Pancreas y Vesicula BiliarEvelina CidОценок пока нет
- Colelitiasis - ColecistitisДокумент7 страницColelitiasis - ColecistitisIziTa AsilemОценок пока нет
- Catálogo Pure Health 2022Документ55 страницCatálogo Pure Health 2022Nancy CarSanОценок пока нет
- HÍGADOДокумент13 страницHÍGADOMariana CaraveoОценок пока нет
- Indicaciones TRX HepaticoДокумент5 страницIndicaciones TRX HepaticoZaira HoriaОценок пока нет
- Practica N°1 de AnatomiaДокумент12 страницPractica N°1 de Anatomiachikitha dayanОценок пока нет
- Sindrome Icterico e Hipertension PortalДокумент8 страницSindrome Icterico e Hipertension PortalMadeline MonteroОценок пока нет
- Wa0000Документ6 страницWa0000Adriana VelascoОценок пока нет
- Antiflatulentos, Antiemeicos y HepaticosДокумент13 страницAntiflatulentos, Antiemeicos y Hepaticoscedelca maicaoОценок пока нет
- Diapositivas Histologia Segundo Parcial Casi Completo-1 PDFДокумент186 страницDiapositivas Histologia Segundo Parcial Casi Completo-1 PDFadriana palizОценок пока нет
- Enzimas HigadoДокумент34 страницыEnzimas HigadoAlejandro GalloОценок пока нет
- Patología VesiculaДокумент10 страницPatología VesiculaFelipe HerreraОценок пока нет
- El Aparato DigestivoДокумент72 страницыEl Aparato DigestivoMore ParedesОценок пока нет
- 02 - Drenaje WebДокумент6 страниц02 - Drenaje WebTANIA PINTO BUSTAMANTEОценок пока нет
- 9a Práctica Lab Anatomia Hígado, Páncreas y Vías BiliaresДокумент2 страницы9a Práctica Lab Anatomia Hígado, Páncreas y Vías BiliaresRuth Cobo RosalesОценок пока нет
- EL HígadoДокумент6 страницEL HígadoAlexa Chinchilla PachecoОценок пока нет
- Wuolah Free Bloque IIIДокумент195 страницWuolah Free Bloque IIIJorge Ocón CeballosОценок пока нет
- Encefalopatía MetabólicaДокумент3 страницыEncefalopatía MetabólicaCarlos Rodrigo SanchezОценок пока нет
- Valoración Radiológica de La Hernia Diafragmática Congénita FetalДокумент9 страницValoración Radiológica de La Hernia Diafragmática Congénita FetalEduardo José Horna GarcíaОценок пока нет
- RX de Tórax: HallazgosДокумент4 страницыRX de Tórax: HallazgosTeofilo CastroОценок пока нет
- NCL2 U1 A2 RipbДокумент10 страницNCL2 U1 A2 RipbSika Bravo100% (1)
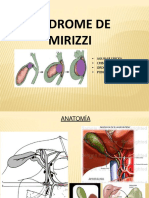
- Sindrome de MirizziДокумент25 страницSindrome de MirizziScribdRandom1100% (1)
- 7 IctericiasparteIДокумент6 страниц7 IctericiasparteIChanego CastilloОценок пока нет
- Colangitis AgudaДокумент4 страницыColangitis Agudacrisllyshantal chuquimamani pariОценок пока нет