Вам также может понравиться
- Bloque 1 Cap 5 Tema 6. Digestion Rumiantes IДокумент5 страницBloque 1 Cap 5 Tema 6. Digestion Rumiantes IClaudio San MartinОценок пока нет
- 12.6 El Aparato Digestivo de La Cabra y La OvejaДокумент5 страниц12.6 El Aparato Digestivo de La Cabra y La OvejaMirella SuarezОценок пока нет
- Anatomia y Fisiologia Del CerdoДокумент10 страницAnatomia y Fisiologia Del CerdoMarta Nuñez100% (2)
- NUTRICIÓN DE RUMIANTES Funcion Del RumenДокумент25 страницNUTRICIÓN DE RUMIANTES Funcion Del RumenJose DeJesus Bustamante GuerreroОценок пока нет
- Digestión y absorción de proteínas en rumiantesДокумент18 страницDigestión y absorción de proteínas en rumiantesMelany BobadillaОценок пока нет
- Motilidad Del RumenДокумент14 страницMotilidad Del RumenSócrates Millman0% (1)
- Anatomía y fisiología del aparato reproductor de la coneja hembraДокумент11 страницAnatomía y fisiología del aparato reproductor de la coneja hembramauricioОценок пока нет
- Cría de Ovinos y CaprinosДокумент8 страницCría de Ovinos y CaprinosNorelis AldanaОценок пока нет
- Sistema Digestivo de CerdosДокумент18 страницSistema Digestivo de CerdosJosé Lóp'zОценок пока нет
- Taller 2 AlimentaciónДокумент4 страницыTaller 2 AlimentaciónCarlos GuerreroОценок пока нет
- Carbohidratos en Nutricion EquinaДокумент5 страницCarbohidratos en Nutricion EquinaAlex D' RoorshackОценок пока нет
- Análisis de la bosta bovina y su relación con la alimentaciónДокумент9 страницAnálisis de la bosta bovina y su relación con la alimentaciónAlain León100% (1)
- Catagña Cristhian - Sistema Renal en Conejos InformeДокумент10 страницCatagña Cristhian - Sistema Renal en Conejos InformeEVELIN DAMARIS QUIROZ TAPIAОценок пока нет
- Aparato Digestivo Del CerdoДокумент8 страницAparato Digestivo Del CerdoJersonLimaymantaSalazarОценок пока нет
- Cavidad Abdominal Del BovinoДокумент10 страницCavidad Abdominal Del BovinoSamanthaVegaОценок пока нет
- Anatomía y Fisiología Del Sistema Digestivo de Aves........Документ5 страницAnatomía y Fisiología Del Sistema Digestivo de Aves........Johan Nicolas QuinteroОценок пока нет
- 0880 Ensayo EquinoДокумент4 страницы0880 Ensayo EquinoLESLY YULIANA BELTRAN GUERRAОценок пока нет
- Sistema Digestivo de Los Animales y Sus DiferenciasДокумент10 страницSistema Digestivo de Los Animales y Sus DiferenciasLuz MataОценок пока нет
- Sistema Digestivo de Animales PoligastricosДокумент6 страницSistema Digestivo de Animales Poligastricoschristina8reales8ca8100% (1)
- Mascotas de Bolsillo. EnsayoДокумент3 страницыMascotas de Bolsillo. EnsayoAndrea KuroNekoОценок пока нет
- BioquДокумент9 страницBioquKatherine Pinduisaca100% (2)
- Alimentación y Nutrición de MonogástricosДокумент18 страницAlimentación y Nutrición de MonogástricosMiguelОценок пока нет
- Nutrición AnimalДокумент9 страницNutrición AnimalYuliana GomezОценок пока нет
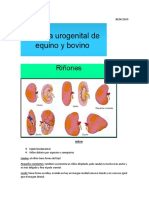
- Sistema UrogenitalДокумент31 страницаSistema UrogenitalSebasthian Alonso Vejar CayupilОценок пока нет
- Abordaje abdominal del BovinoДокумент43 страницыAbordaje abdominal del BovinoLaura Plata33% (3)
- Proyecto Ing. MONTAÑOДокумент10 страницProyecto Ing. MONTAÑORichard Suárez VelardeОценок пока нет
- Músculos Del Cuello PDFДокумент27 страницMúsculos Del Cuello PDFJesenia Segura0% (1)
- Urolitiasis en OvinosДокумент14 страницUrolitiasis en Ovinosomarzito_mexflow5041100% (2)
- Anatomia y Fisiologia de Los RumiantesДокумент37 страницAnatomia y Fisiologia de Los Rumiantesdaddy rey0% (1)
- Ganado bovino carneДокумент10 страницGanado bovino carneAlejandro AguilarОценок пока нет
- El Sistema Digestivo MonogastricoДокумент3 страницыEl Sistema Digestivo MonogastricoBryan-OcrasОценок пока нет
- APO 13 Lagomorfos y Roedores NegraДокумент45 страницAPO 13 Lagomorfos y Roedores NegraMarieОценок пока нет
- Cavidad Nasal PDFДокумент37 страницCavidad Nasal PDFMirta Graciela Sack100% (1)
- Aparato Sustentador Del Equino PDFДокумент3 страницыAparato Sustentador Del Equino PDFErika Adriana S. RojasОценок пока нет
- Acidosis RuminalДокумент2 страницыAcidosis RuminalPablo SalazarОценок пока нет
- EQUINOSДокумент101 страницаEQUINOSIngrid EstradaОценок пока нет
- Aparato digestivo equino: estómago simple, intestino delgado de 22-24m e intestino grueso con ciego de 1-1.2mДокумент2 страницыAparato digestivo equino: estómago simple, intestino delgado de 22-24m e intestino grueso con ciego de 1-1.2mNancy Yasmin PinedaОценок пока нет
- La Palpacion Rectal en Bovinos para Diagnostico Reproductivo 2019Документ16 страницLa Palpacion Rectal en Bovinos para Diagnostico Reproductivo 2019Daniel Flores PérezОценок пока нет
- ConejosДокумент24 страницыConejosSilvana LindaОценок пока нет
- Proceso de La RumiaДокумент2 страницыProceso de La RumiaenriqueОценок пока нет
- Miología Del EquinoДокумент18 страницMiología Del EquinoAntony RafaelОценок пока нет
- Sistema Digestivo Del CerdoДокумент13 страницSistema Digestivo Del CerdoleonardoОценок пока нет
- Fisiología Digestiva de Los AnimalesДокумент16 страницFisiología Digestiva de Los AnimalesJoel Gonzalez PittyОценок пока нет
- Anaplasmosis y BabesiosisДокумент29 страницAnaplasmosis y BabesiosisKatty ZanabriaОценок пока нет
- Sistema digestivo conejosДокумент6 страницSistema digestivo conejosalfre mayОценок пока нет
- Sistema Digestivo EquinoДокумент11 страницSistema Digestivo EquinoAndrea PerezОценок пока нет
- PDF document-20342CD7507B-1Документ35 страницPDF document-20342CD7507B-1Antonio NuñezОценок пока нет
- Fisiología Digestiva en RumiantesДокумент19 страницFisiología Digestiva en RumiantesPablo Cesar Requena NeyraОценок пока нет
- Sistema Digestivo RumianteДокумент11 страницSistema Digestivo Rumiantejorge eliecer carrillo romeroОценок пока нет
- Sistema Digestivo de Los EquinosДокумент26 страницSistema Digestivo de Los EquinosMayra MarcallaОценок пока нет
- Fisiología PorcinaДокумент19 страницFisiología PorcinaFrancoОценок пока нет
- Manual de Sanidad Animal OfiДокумент14 страницManual de Sanidad Animal OfiIngriid VillänueväОценок пока нет
- Trabajo de Sistemas de Pastoreo 1Документ7 страницTrabajo de Sistemas de Pastoreo 1Wveimar Mon CarОценок пока нет
- Aparato Reproductor de La MarranaДокумент4 страницыAparato Reproductor de La MarranaRider SantiagoОценок пока нет
- Hibridación de CamélidosДокумент29 страницHibridación de CamélidosLuis Humberto Inga HuertasОценок пока нет
- JuzgamientoДокумент101 страницаJuzgamientoSofy FranОценок пока нет
- Sistema Urinario-ComparadaДокумент14 страницSistema Urinario-ComparadaCharles Rodriguez100% (1)
- Leguminosas espontaneas de posible valor forrajero en ColombiaОт EverandLeguminosas espontaneas de posible valor forrajero en ColombiaОценок пока нет
- Instalaciones, maquinaria y equipos de la explotación ganadera. AGAP0108От EverandInstalaciones, maquinaria y equipos de la explotación ganadera. AGAP0108Оценок пока нет
- Semana 15 - Uso y Aprovechamiento de La FloraДокумент20 страницSemana 15 - Uso y Aprovechamiento de La FloraEnrique ContrerasОценок пока нет
- 1.trabajo de EmpaqueДокумент13 страниц1.trabajo de EmpaqueSotomayor ReinaldoОценок пока нет
- ColibrisДокумент4 страницыColibrismartacrodriguezОценок пока нет
- Exposicion Region InsularДокумент5 страницExposicion Region InsularllpaОценок пока нет
- Estas Son Las Mejores Cervezas Lager de Marca Blanca Según El Catador Carlos Gómez - Noticias deДокумент4 страницыEstas Son Las Mejores Cervezas Lager de Marca Blanca Según El Catador Carlos Gómez - Noticias deJorge Olivares HuertasОценок пока нет
- Cadena de Abast ALPINAДокумент15 страницCadena de Abast ALPINAAndres Ortega50% (2)
- BPM Carne de CerdoДокумент383 страницыBPM Carne de CerdoGina Nuñez100% (5)
- Reactivos Gestion de La CalidadДокумент7 страницReactivos Gestion de La CalidadKevin MopoОценок пока нет
- Mosquita Blanca PDFДокумент4 страницыMosquita Blanca PDFFernandoОценок пока нет
- Costa o ChalaДокумент17 страницCosta o ChalaLuisa Leen100% (3)
- #5 Indicadores de Educacion, Salud y Vivienda Del PeruДокумент18 страниц#5 Indicadores de Educacion, Salud y Vivienda Del PeruPablo RuizОценок пока нет
- Trabajo de PericoДокумент31 страницаTrabajo de PericoEmily Lucía JZОценок пока нет
- Órganos Zang Fu 2012Документ33 страницыÓrganos Zang Fu 2012Oscar FloresОценок пока нет
- Conteo de CarbohidratosДокумент79 страницConteo de Carbohidratoscarlos enriqueОценок пока нет
- Guia AgroindustriaДокумент100 страницGuia AgroindustriaCesar Yvan Facundo PuellesОценок пока нет
- Actividades de Enfermería para La Detección OportunaДокумент4 страницыActividades de Enfermería para La Detección OportunaCarlos Omar TapiaОценок пока нет
- Batidos saludables para desintoxicar y perder grasaДокумент13 страницBatidos saludables para desintoxicar y perder grasaVarinia Salazar SmithОценок пока нет
- Ecoswing - HTДокумент3 страницыEcoswing - HTAlexОценок пока нет
- Parcial Teoría de Colas - FinalДокумент6 страницParcial Teoría de Colas - FinalAngela Sanchez MartinezОценок пока нет
- La Presentación de La Empresa ALPINAДокумент10 страницLa Presentación de La Empresa ALPINAKelly DiaZОценок пока нет
- LA EXCUSA - La Culpa Es de La VacaДокумент7 страницLA EXCUSA - La Culpa Es de La VacaGuillermo AlfaraОценок пока нет
- Caso de Estudio - Retail ManagementДокумент35 страницCaso de Estudio - Retail ManagementSeminario Seccion 1Оценок пока нет
- Informe de Administración de La Cadena de Suministro en MC DonaldsДокумент6 страницInforme de Administración de La Cadena de Suministro en MC DonaldsJuliana María Amaya PinillaОценок пока нет
- Estudio Calidad Galletas Marias Saladas y AnimalitosДокумент25 страницEstudio Calidad Galletas Marias Saladas y AnimalitosArelyОценок пока нет
- Diagrama de Flujo para La Elaboración Del Vino Espumoso de MandarinaДокумент6 страницDiagrama de Flujo para La Elaboración Del Vino Espumoso de MandarinaTahic P0% (1)
- Alimentacion de GallinasДокумент9 страницAlimentacion de GallinasJarol Andres Hoyos JoaquiОценок пока нет
- Alimentación A Niños Menores de 2 Años CreoleДокумент32 страницыAlimentación A Niños Menores de 2 Años CreoleMaría Belén DíazОценок пока нет
- Unidad 1 Ciencias NaturalesДокумент17 страницUnidad 1 Ciencias NaturalesDimmy Jael Victoria Moya QuirozОценок пока нет
- Ingredientes cerveza artesanalДокумент3 страницыIngredientes cerveza artesanalJuan David Rodriguez EspitiaОценок пока нет
- Recetas de Pan CaseroДокумент26 страницRecetas de Pan CaseroPepe TejasОценок пока нет