Вам также может понравиться
- Plant Resistance to Herbivores and Pathogens: Ecology, Evolution, and GeneticsОт EverandPlant Resistance to Herbivores and Pathogens: Ecology, Evolution, and GeneticsОценок пока нет
- Ecological GeneticДокумент8 страницEcological GeneticMickaël Houessou SononОценок пока нет
- Áfidos y MonarcasДокумент9 страницÁfidos y MonarcasMario RestrepoОценок пока нет
- Jones 1999Документ3 страницыJones 1999Md. Sohanur RahmanОценок пока нет
- New Phytologist - 2013 - Shahzad - Plant Defensin Type 1 PDF1 Protein Promiscuity and Expression Variation Within TheДокумент14 страницNew Phytologist - 2013 - Shahzad - Plant Defensin Type 1 PDF1 Protein Promiscuity and Expression Variation Within Thedora guzmanОценок пока нет
- Ecological and Population Genetics of Herbicide Resistance: Where To Now?Документ4 страницыEcological and Population Genetics of Herbicide Resistance: Where To Now?Esteban Muñiz PadillaОценок пока нет
- International Journal of BiologyДокумент26 страницInternational Journal of BiologyAnnisa Khoirun Nihayah almalikiОценок пока нет
- Marinedrugs 05 00220Документ22 страницыMarinedrugs 05 00220Muhammad Satria AldinoОценок пока нет
- Tipping Competition Aquatic PlantsДокумент4 страницыTipping Competition Aquatic PlantsLeonardo Daniel OrtegaОценок пока нет
- Both Plant Genotype and Herbivory Shape Aspen Endophyte CommunitiesДокумент11 страницBoth Plant Genotype and Herbivory Shape Aspen Endophyte CommunitiesgkcnbpvyiyjwotryjaОценок пока нет
- Introduction NewДокумент12 страницIntroduction NewYOHANNIS SHAFEОценок пока нет
- Coprophilous Fungi Antibiotic DiscoveryДокумент17 страницCoprophilous Fungi Antibiotic DiscoveryNathan Ulises Reyes JimenezОценок пока нет
- Chapter 4 Beneficial Arthropods and MicroorganismsДокумент41 страницаChapter 4 Beneficial Arthropods and MicroorganismsDennMark CentenoОценок пока нет
- Genome-Based Evolutionary History of Pseudomonas SPPДокумент51 страницаGenome-Based Evolutionary History of Pseudomonas SPPnfernandezОценок пока нет
- tmpCE23 TMPДокумент14 страницtmpCE23 TMPFrontiersОценок пока нет
- Terpenoids in Plant and Arbuscular Mycorrhiza-Reinforced Defence Against Herbivorous InsectsДокумент11 страницTerpenoids in Plant and Arbuscular Mycorrhiza-Reinforced Defence Against Herbivorous InsectsyogiОценок пока нет
- Mapping of Cotton-Melon Aphid Resistance in MelonДокумент8 страницMapping of Cotton-Melon Aphid Resistance in MelonanradeljaОценок пока нет
- Effects of Resource Distribution on Animal Plant InteractionsОт EverandEffects of Resource Distribution on Animal Plant InteractionsОценок пока нет
- Parasitic Plants: Parallels and Contrasts With Herbivores: Steven C. Pennings Ragan M. CallawayДокумент11 страницParasitic Plants: Parallels and Contrasts With Herbivores: Steven C. Pennings Ragan M. CallawayAthena NocetoОценок пока нет
- 1996 Bever EsporulacaoДокумент13 страниц1996 Bever EsporulacaoMaristela VolpatoОценок пока нет
- Arnold, Lutzoni - 2007 - Diversity and Host Range of Foliar Fungal Endophytes Are Tropical Leaves Biodiversity HotspotsДокумент9 страницArnold, Lutzoni - 2007 - Diversity and Host Range of Foliar Fungal Endophytes Are Tropical Leaves Biodiversity HotspotsLavinia BarbuОценок пока нет
- Evolution 2014 Gibson AKДокумент28 страницEvolution 2014 Gibson AKAlfredo PerettiОценок пока нет
- Density-Dependent Prophylaxis in The Mealworm Beetle Tenebrio Molitor L. (Coleoptera: Tenebrionidae) : Cuticular Melanization Is An Indicator of Investment in ImmunityДокумент6 страницDensity-Dependent Prophylaxis in The Mealworm Beetle Tenebrio Molitor L. (Coleoptera: Tenebrionidae) : Cuticular Melanization Is An Indicator of Investment in ImmunityajudОценок пока нет
- Phytochemistry: Nicol As Cara, Patricia N. Piccoli, Leonardo Bolcato, Carlos F. Marfil, Ricardo W. MasuelliДокумент8 страницPhytochemistry: Nicol As Cara, Patricia N. Piccoli, Leonardo Bolcato, Carlos F. Marfil, Ricardo W. MasuelliLuisОценок пока нет
- Bacterial Endophytes: Recent Developments and ApplicationsДокумент9 страницBacterial Endophytes: Recent Developments and ApplicationsmaczweiОценок пока нет
- M V. C S A. K: Orrichia Frutescens and - ArborescensДокумент10 страницM V. C S A. K: Orrichia Frutescens and - Arborescensnica_salvadorОценок пока нет
- Friends or Foes? Emerging Insights From Fungal Interactions With PlantsДокумент26 страницFriends or Foes? Emerging Insights From Fungal Interactions With PlantsSandii Achmad ApriliantoОценок пока нет
- M YCROZIHAДокумент63 страницыM YCROZIHASamuel DavisОценок пока нет
- Chapter1 Howe Schaller 01Документ23 страницыChapter1 Howe Schaller 01Anushka SujeewonОценок пока нет
- Discussion - Final OoДокумент3 страницыDiscussion - Final OofupaОценок пока нет
- Welcome: Topic To Be PresentedДокумент29 страницWelcome: Topic To Be PresentedChiranjit DebbarmaОценок пока нет
- CBO9780511902451A015Документ20 страницCBO9780511902451A015Veronica B MarinaОценок пока нет
- 2011 Rojas Et Al Plantas Filostomidos PDFДокумент12 страниц2011 Rojas Et Al Plantas Filostomidos PDFJ E Fernando CernaОценок пока нет
- Diversity and Environmental Stress Responses of Rhizobial Bacteria From Egyptian Grain LegumesДокумент13 страницDiversity and Environmental Stress Responses of Rhizobial Bacteria From Egyptian Grain LegumesAlbert MangopangОценок пока нет
- Biology Letters - Community Ecology (Bardgett)Документ5 страницBiology Letters - Community Ecology (Bardgett)PololoОценок пока нет
- 3027 FullДокумент12 страниц3027 FullGebrehiwot GebremedhinОценок пока нет
- Paper 4 (Kel 4) (BA4107)Документ9 страницPaper 4 (Kel 4) (BA4107)Fikri SabilalОценок пока нет
- TMP DA00Документ10 страницTMP DA00FrontiersОценок пока нет
- Endophytes To The Rescue of Plants!: Bhavdish N. JohriДокумент2 страницыEndophytes To The Rescue of Plants!: Bhavdish N. JohriLuzAvОценок пока нет
- Phylogenetic and Phytochemical Characteristics Ch8 KLEINДокумент22 страницыPhylogenetic and Phytochemical Characteristics Ch8 KLEINrobynkleinОценок пока нет
- Catatan VilepДокумент2 страницыCatatan VilepaminahtunОценок пока нет
- Ajb 89 3 472Документ7 страницAjb 89 3 472Kesia ParambanОценок пока нет
- SherametietalДокумент24 страницыSherametietalErjon MamociОценок пока нет
- pb16 4 255Документ7 страницpb16 4 255rishee123Оценок пока нет
- Sanchez Et Al 2010Документ12 страницSanchez Et Al 2010minervaОценок пока нет
- 1a82e3bb25d3d7dfd4545296161a090cДокумент40 страниц1a82e3bb25d3d7dfd4545296161a090cmuhammad hamzaОценок пока нет
- Widespread Whole Genome DuplicationsДокумент15 страницWidespread Whole Genome DuplicationssalarОценок пока нет
- Understanding The Diversity of Foliar Endophytic Fungi: Progress, Challenges, and FrontiersДокумент16 страницUnderstanding The Diversity of Foliar Endophytic Fungi: Progress, Challenges, and FrontiersNancy HutchinsonОценок пока нет
- 2007 Phylogenetic and Ohylogenimis Definitions of Rhizopus SpeciesДокумент12 страниц2007 Phylogenetic and Ohylogenimis Definitions of Rhizopus SpeciesPierrotОценок пока нет
- Ecology of Bacterial Endophytes in Sustainable Agriculture: Pablo R. Hardoim, Riitta Nissinen, and Jan D. Van ElsasДокумент32 страницыEcology of Bacterial Endophytes in Sustainable Agriculture: Pablo R. Hardoim, Riitta Nissinen, and Jan D. Van ElsasGregorio AroneОценок пока нет
- 1 s2.0 S0038071721000213 MainДокумент7 страниц1 s2.0 S0038071721000213 MainEmilio VarColОценок пока нет
- Comparison of Community Composition of ParasitoidsДокумент10 страницComparison of Community Composition of ParasitoidsAnonymous Vs4p938sTОценок пока нет
- Production of Bioproducts by Endophytic Fungi: Chemical Ecology, Biotechnological Applications, Bottlenecks, and SolutionsДокумент20 страницProduction of Bioproducts by Endophytic Fungi: Chemical Ecology, Biotechnological Applications, Bottlenecks, and SolutionsScarllett Lalesca Santos LimaОценок пока нет
- Mpmi 05 11 0116Документ11 страницMpmi 05 11 0116Mhd Hady NugrohoОценок пока нет
- Impact of Antifungals Producing Rhizobacteria On The Performance of Vigna Radiata in The Presence of Arbuscular Mycorrhizal FungiДокумент12 страницImpact of Antifungals Producing Rhizobacteria On The Performance of Vigna Radiata in The Presence of Arbuscular Mycorrhizal FungimetbijayОценок пока нет
- Carmona Et Al. 2013 - Acta OecologicaДокумент7 страницCarmona Et Al. 2013 - Acta OecologicaCarlos Pérez CarmonaОценок пока нет
- Phytochemical Variation On Tree Tops 2018 LamkeДокумент12 страницPhytochemical Variation On Tree Tops 2018 LamkeClaude MenodzОценок пока нет
- Molecular and Cell Biology of Arbuscular Mycorrhizal SymbiosisДокумент13 страницMolecular and Cell Biology of Arbuscular Mycorrhizal SymbiosisGiorgio PredelliОценок пока нет
- Whipps Et Al, 2008Документ12 страницWhipps Et Al, 2008joyeeta8Оценок пока нет
- Day 9 - Training Business WritingДокумент23 страницыDay 9 - Training Business WritingArreza Hamsyah FernandaОценок пока нет
- 50 Liquid Citric AcidДокумент8 страниц50 Liquid Citric AcidArreza Hamsyah FernandaОценок пока нет
- Master Harga SaraswantiДокумент12 страницMaster Harga SaraswantiArreza Hamsyah FernandaОценок пока нет
- UASДокумент6 страницUASArreza Hamsyah FernandaОценок пока нет
- Nick de Vries - Motivation Letter Junior Consultant-1 PDFДокумент1 страницаNick de Vries - Motivation Letter Junior Consultant-1 PDFArreza Hamsyah FernandaОценок пока нет
- MicrologДокумент1 страницаMicrologArreza Hamsyah FernandaОценок пока нет
- 23 45 1 SM PDFДокумент7 страниц23 45 1 SM PDFArreza Hamsyah FernandaОценок пока нет
- Ultra Low Temperature Freezer LabДокумент2 страницыUltra Low Temperature Freezer LabArreza Hamsyah FernandaОценок пока нет
- License PremiumДокумент1 страницаLicense PremiumNguyen Viet Trung (FPL HCMK13.3)100% (1)
- Industrial Biotechnology 8Документ35 страницIndustrial Biotechnology 8Arreza Hamsyah FernandaОценок пока нет
- Hi Arreza HF,: Thank You For Using GO-RIDE, We Hope You Enjoyed Riding With UsДокумент1 страницаHi Arreza HF,: Thank You For Using GO-RIDE, We Hope You Enjoyed Riding With UsArreza Hamsyah FernandaОценок пока нет
- Presentation 1Документ17 страницPresentation 1Arreza Hamsyah FernandaОценок пока нет
- Price List HondaДокумент1 страницаPrice List HondaArreza Hamsyah FernandaОценок пока нет
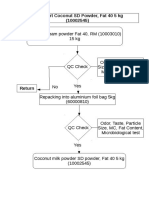
- Flowchart Coconut SD Powder, Fat 40 5 KG (10002545)Документ1 страницаFlowchart Coconut SD Powder, Fat 40 5 KG (10002545)Arreza Hamsyah FernandaОценок пока нет
- Uji Sterilitas FarmakopeДокумент0 страницUji Sterilitas FarmakopeEkaekaekaaОценок пока нет
- Flowchart Coconut SD Powder, Fat 40 5 KG (10002545)Документ1 страницаFlowchart Coconut SD Powder, Fat 40 5 KG (10002545)Arreza Hamsyah FernandaОценок пока нет
- Ceramic Fiber 23Документ28 страницCeramic Fiber 23Kowsik RajendranОценок пока нет
- Unit Weight of Materials Used at Construction Site - Online Civil PDFДокумент2 страницыUnit Weight of Materials Used at Construction Site - Online Civil PDFKeshav VarpeОценок пока нет
- RNA and Protein Synthesis Problem SetДокумент6 страницRNA and Protein Synthesis Problem Setpalms thatshatterОценок пока нет
- Daftar Irregular VerbsДокумент8 страницDaftar Irregular VerbsWawan MarwanОценок пока нет
- 1896 - Pearson - Mathematical Contributions To The Theory of Evolution. III. Regression, Heredity, and PanmixiaДокумент67 страниц1896 - Pearson - Mathematical Contributions To The Theory of Evolution. III. Regression, Heredity, and PanmixiaNilotpal N SvetlanaОценок пока нет
- Master Handover Rev 4-2013Документ8 страницMaster Handover Rev 4-2013MeysamGholizadehОценок пока нет
- BFPPPДокумент15 страницBFPPPFaith JacalanОценок пока нет
- ThreeДокумент6 страницThreeapi-340425056Оценок пока нет
- Chapter 8 Sensation and PerceptionДокумент66 страницChapter 8 Sensation and Perceptionapi-726122866Оценок пока нет
- Am Jf211 - Jul 04Документ4 страницыAm Jf211 - Jul 04ilham_metallurgy6744Оценок пока нет
- Trump ReportДокумент237 страницTrump ReportAaron Nobel100% (1)
- SCOPE OF PRACTICE FOR TCAM PRACTITONERS - V - 01Документ22 страницыSCOPE OF PRACTICE FOR TCAM PRACTITONERS - V - 01shakkiryousufОценок пока нет
- Daftar Pustaka ProposalДокумент4 страницыDaftar Pustaka ProposalraniОценок пока нет
- Drugstudy - Delivery RoomДокумент12 страницDrugstudy - Delivery RoomAUBREY MARIE . GUERREROОценок пока нет
- HEM Duplex Cooling MotorДокумент4 страницыHEM Duplex Cooling MotorAbhishek Kumar Pandey1Оценок пока нет
- vdYoyHdeTKeL7EhJwoXE - Insomnia PH SlidesДокумент40 страницvdYoyHdeTKeL7EhJwoXE - Insomnia PH SlidesKreshnik IdrizajОценок пока нет
- Construction Quality Control PlanДокумент35 страницConstruction Quality Control PlanMalik Riaz100% (4)
- Aqa Chm6w W QP Jun02Документ18 страницAqa Chm6w W QP Jun02Diksha KoossoolОценок пока нет
- Football Speed TrainingДокумент6 страницFootball Speed TrainingTBM607Оценок пока нет
- Flexible Learnin G: Group 3 Bsed-Math 2Документ48 страницFlexible Learnin G: Group 3 Bsed-Math 2Niña Gel Gomez AparecioОценок пока нет
- Acute GastroenteritisДокумент7 страницAcute GastroenteritisCherr NollОценок пока нет
- Pipe Conveyor System: Effective Solutions For Long DistancesДокумент8 страницPipe Conveyor System: Effective Solutions For Long DistancesEugenepaccelli Kolandai SamyОценок пока нет
- Plumbing Engineering Design Handbook, Vol 1 ASPEДокумент422 страницыPlumbing Engineering Design Handbook, Vol 1 ASPEMichelle NataliОценок пока нет
- Review of Documents On Seismic Strengthening of Existing BuildingsДокумент12 страницReview of Documents On Seismic Strengthening of Existing Buildingsm7j7a7Оценок пока нет
- Cde-Class II Amalgam Restorations-31!12!14Документ36 страницCde-Class II Amalgam Restorations-31!12!14Archita KureelОценок пока нет
- Devoir de Contrôle N°2 - Anglais - 2ème Economie & Gestion (2015-2016) Mme Chaima JallebДокумент3 страницыDevoir de Contrôle N°2 - Anglais - 2ème Economie & Gestion (2015-2016) Mme Chaima JallebSassi LassaadОценок пока нет
- Steri - Cycle I 160 New GenДокумент16 страницSteri - Cycle I 160 New GenLEO AROKYA DASSОценок пока нет
- Strip Seal Expansion JointДокумент13 страницStrip Seal Expansion JointsmithОценок пока нет
- Personal Loan For JoiningДокумент3 страницыPersonal Loan For JoiningAzim SengalОценок пока нет
- Issues Pertaining To Maintenance of WifeДокумент2 страницыIssues Pertaining To Maintenance of WifeVaishnavi YadavОценок пока нет