Вам также может понравиться
- Influencia de Las Acuaporinas en La Génesis de Múltiples EnfermedadesДокумент9 страницInfluencia de Las Acuaporinas en La Génesis de Múltiples EnfermedadesFernando Rios ZeladaОценок пока нет
- Acuaporinas PDFДокумент5 страницAcuaporinas PDFKevin NietoОценок пока нет
- Informe de SeminarioДокумент6 страницInforme de SeminarioJASON ALBERTO VILCAHUAMAN PUCHOCОценок пока нет
- Anexo 3 Biologia Aquaporinas 1Документ5 страницAnexo 3 Biologia Aquaporinas 1Aldana GonzalezОценок пока нет
- Acuaporinas y Edema CerebralДокумент10 страницAcuaporinas y Edema CerebralAnthony Christian Rivera SánchezОценок пока нет
- Estructura, Función y Localización de Las AcuaporinasДокумент36 страницEstructura, Función y Localización de Las AcuaporinasAugusto Tamani100% (1)
- Canales acuaporinasДокумент8 страницCanales acuaporinasDiego IgnacioОценок пока нет
- ACUAPORINAS: Proteínas clave en el transporte de aguaДокумент2 страницыACUAPORINAS: Proteínas clave en el transporte de aguaNestor Raul SotoОценок пока нет
- Influencia de Las Acuaporinas en La Génesis de Múltiples Enfermedades - 2014Документ15 страницInfluencia de Las Acuaporinas en La Génesis de Múltiples Enfermedades - 2014MariaGladisCorderoVallejoОценок пока нет
- Resumen AcuaporinasДокумент4 страницыResumen AcuaporinasMichelle Acuña EspinozaОценок пока нет
- AcuaporinaДокумент8 страницAcuaporinakmel09Оценок пока нет
- Articulo AcuaporinasДокумент12 страницArticulo AcuaporinasKatherine Pradilla ZambranoОценок пока нет
- Acuaporina LameckДокумент11 страницAcuaporina LamecklamecОценок пока нет
- Cuestionario de AcuaporinasДокумент8 страницCuestionario de AcuaporinasYair Arriaga KanahanОценок пока нет
- AcuaporinasДокумент5 страницAcuaporinasJuan Jose DiazОценок пока нет
- AcuaporinasДокумент12 страницAcuaporinasPeter P. R. MoralesОценок пока нет
- Acuaporina Canales de AguaДокумент3 страницыAcuaporina Canales de AguaDesireé Colio MedinaОценок пока нет
- Acu A PorinasДокумент3 страницыAcu A PorinasAlieska Salazar CabreraОценок пока нет
- Acua PorinasДокумент9 страницAcua PorinasdarielОценок пока нет
- 01 Echevarría M & R Zardoya (2006) AcuaporinasДокумент8 страниц01 Echevarría M & R Zardoya (2006) AcuaporinasCristobal Aguila RebolledoОценок пока нет
- AcuaporinaДокумент8 страницAcuaporinaRicardo FelixОценок пока нет
- Acuaporinas en El Sistema RespiratorioДокумент3 страницыAcuaporinas en El Sistema RespiratorioJoselyNoemiSánchezMedina0% (2)
- AcuaporinasДокумент3 страницыAcuaporinasrubipavia21Оценок пока нет
- De La Estructura A La EnfermedadДокумент4 страницыDe La Estructura A La Enfermedadapi-3807078100% (1)
- Las AcuaporinasДокумент9 страницLas AcuaporinasNaiara Alonso VilchesОценок пока нет
- ACUAPORINAДокумент2 страницыACUAPORINARodОценок пока нет
- Acuaporinas en plantas y su posible rol en la regulación del intercambio de agua y CO2 en HymenophyllaceasДокумент30 страницAcuaporinas en plantas y su posible rol en la regulación del intercambio de agua y CO2 en Hymenophyllaceasnnnanda39Оценок пока нет
- Acuaporina - Wikipedia, La Enciclopedia LibreДокумент9 страницAcuaporina - Wikipedia, La Enciclopedia LibreLorenzo JimenezОценок пока нет
- Canales de agua celulares: Las acuaporinas y su descubrimientoДокумент30 страницCanales de agua celulares: Las acuaporinas y su descubrimientoZoila Paredes100% (1)
- Acuaporinas: Proteínas Mediadoras Del Transporte de Agua.Документ8 страницAcuaporinas: Proteínas Mediadoras Del Transporte de Agua.Nathalia Acevedo EscalanteОценок пока нет
- Título Del Ensayo - AcuaporinasДокумент2 страницыTítulo Del Ensayo - AcuaporinasPëlaïthö Gualli KevinОценок пока нет
- Acuaporinas en El Sistema RespiratorioДокумент2 страницыAcuaporinas en El Sistema RespiratorioMiguel ChavezОценок пока нет
- Acu A PorinasДокумент5 страницAcu A PorinasChristian Fernando Rodriguez JimenesОценок пока нет
- AQUAPORINASДокумент5 страницAQUAPORINASMary Bernal SaraviaОценок пока нет
- Qué Son Las AcuaporinasДокумент2 страницыQué Son Las AcuaporinasDaniela ayalaОценок пока нет
- Las Acuaporinas y Agua La Homeostasis en Las PlantasДокумент18 страницLas Acuaporinas y Agua La Homeostasis en Las PlantasRaul Barja LorenzoОценок пока нет
- Aguaporina ExposicionДокумент14 страницAguaporina ExposicionIngrid ThomasОценок пока нет
- Taller de LecturaДокумент2 страницыTaller de LecturaruizaguilarmonicaabigailОценок пока нет
- Fisiologia Humana AcuaporinaДокумент3 страницыFisiologia Humana AcuaporinaJose MachucaОценок пока нет
- Lectura 2 Membrana Celular y AcuaporinasДокумент6 страницLectura 2 Membrana Celular y AcuaporinasTenazОценок пока нет
- Canales acuosos celulares: descubrimiento de las acuaporinasДокумент8 страницCanales acuosos celulares: descubrimiento de las acuaporinasjavieraОценок пока нет
- AquaporinasДокумент21 страницаAquaporinasNahumОценок пока нет
- Acuaporinas: Estructura, Función y ClasificaciónДокумент3 страницыAcuaporinas: Estructura, Función y ClasificaciónCarlos HerreraОценок пока нет
- Regulación de La Expresión de Acuaporinas Por Hormonas OváricasДокумент3 страницыRegulación de La Expresión de Acuaporinas Por Hormonas OváricasDenisse ZayasОценок пока нет
- Acu A PorinasДокумент45 страницAcu A Porinasbrunocs10Оценок пока нет
- Acuaporinas: canales de agua celulares(40Документ2 страницыAcuaporinas: canales de agua celulares(40eliocesar92Оценок пока нет
- Dónde Se Produce Las ProstaglandinasДокумент5 страницDónde Se Produce Las ProstaglandinasLEIDY SALOME CUENCA CELIОценок пока нет
- Informe de Bio 8Документ4 страницыInforme de Bio 8ccalleca01Оценок пока нет
- Acuaporinas A-1 PDFДокумент4 страницыAcuaporinas A-1 PDFCindy Yudith Arotoma LeguiaОценок пока нет
- Fisiología hepática y pancreáticaДокумент31 страницаFisiología hepática y pancreáticaLorenaОценок пока нет
- AQUAPORINA 8- BIOQUÍMICA-24 hoy kathia[1] AVANCE HOY[1]Документ4 страницыAQUAPORINA 8- BIOQUÍMICA-24 hoy kathia[1] AVANCE HOY[1]tik tok videosОценок пока нет
- AQUAPORINA 8- BIOQUÍMICA-24 hoy kathia[1]Документ3 страницыAQUAPORINA 8- BIOQUÍMICA-24 hoy kathia[1]tik tok videosОценок пока нет
- Informe Motilidad Gastrointestinal Del Cuy Fisio2Документ9 страницInforme Motilidad Gastrointestinal Del Cuy Fisio2Nilo Choquehuanca ChoquenairaОценок пока нет
- Fisiopatología de La Pancreatitis Aguda: Revision de Temas CirugiaДокумент8 страницFisiopatología de La Pancreatitis Aguda: Revision de Temas CirugiaJonathan PoloОценок пока нет
- El puzle del agua y la clave exagonal: Evidencias científicas del Agua Hexagonal y su influencia en la vidaОт EverandEl puzle del agua y la clave exagonal: Evidencias científicas del Agua Hexagonal y su influencia en la vidaРейтинг: 5 из 5 звезд5/5 (1)
- Rejuvenecer Con El Plasma Sanguíneo De Los JóvenesОт EverandRejuvenecer Con El Plasma Sanguíneo De Los JóvenesРейтинг: 5 из 5 звезд5/5 (1)
- Metabolismo del sistema digestivo, del hígado, de la vesícula y de las vías biliares: En condiciones de salud y en las enfermedadesОт EverandMetabolismo del sistema digestivo, del hígado, de la vesícula y de las vías biliares: En condiciones de salud y en las enfermedadesОценок пока нет
- Fisio1. Sem3 Fisio Celular. Práctica. Solutos y Agua. (EDA+shock Hipovolemico. Falla Cardiaca Por Valvulopatia Post Reumatica) PDFДокумент1 страницаFisio1. Sem3 Fisio Celular. Práctica. Solutos y Agua. (EDA+shock Hipovolemico. Falla Cardiaca Por Valvulopatia Post Reumatica) PDFDr. Harry'SОценок пока нет
- Fisio1. Sem2 Fisio Celular. Seminario. Transducción de Señales PDFДокумент1 страницаFisio1. Sem2 Fisio Celular. Seminario. Transducción de Señales PDFDr. Harry'SОценок пока нет
- Fisio1. Sem1 Fisio Celular. Práctica. Casos Clínicos PDFДокумент2 страницыFisio1. Sem1 Fisio Celular. Práctica. Casos Clínicos PDFDr. Harry'SОценок пока нет
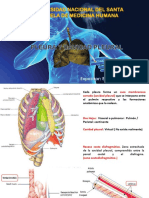
- Anatomia Exposicion PleuraДокумент12 страницAnatomia Exposicion PleuraDr. Harry'SОценок пока нет
- AcuaporinasДокумент8 страницAcuaporinasFabiola MartinezОценок пока нет
- Los Grandes Mecanismos Disipativos y Sus Fuerzas ImpulsorasДокумент14 страницLos Grandes Mecanismos Disipativos y Sus Fuerzas ImpulsorasCoti Allovero100% (1)
- 3 Mecanismos de TransporteДокумент54 страницы3 Mecanismos de TransporteBianca PercsiОценок пока нет
- Resumen Capítulo 4 Transporte de Sustancias A Través de Las Membranas CelularesДокумент10 страницResumen Capítulo 4 Transporte de Sustancias A Través de Las Membranas CelularesJhonatan Vasquez RojasОценок пока нет
- 9no Teo TRANSPORTE 2 PARTE 2Документ5 страниц9no Teo TRANSPORTE 2 PARTE 2Florencia PonceОценок пока нет
- Acuaporines1 1Документ27 страницAcuaporines1 1Frida ToledoОценок пока нет
- SEMINARIO 3 UsmpДокумент14 страницSEMINARIO 3 UsmpHenry Alfredo Vargas RojasОценок пока нет
- Acuaporinas A-1 PDFДокумент4 страницыAcuaporinas A-1 PDFCindy Yudith Arotoma LeguiaОценок пока нет
- Parte 4-Membrana y Mecanismos de TransporteДокумент52 страницыParte 4-Membrana y Mecanismos de TransporteDalia ChavarriaОценок пока нет
- Estructura AcuaporinasДокумент8 страницEstructura AcuaporinasERIK VÉLEZОценок пока нет
- Capítulo 5 - Membranas BiológicasДокумент28 страницCapítulo 5 - Membranas Biológicassan0423Оценок пока нет
- TEMA 4 ImagesДокумент99 страницTEMA 4 Imagesismael.laasikriОценок пока нет
- Tema 3. Potencial HídricoДокумент8 страницTema 3. Potencial HídricoJosé Álvarez del HoyoОценок пока нет
- Análisis rápido sueloДокумент16 страницAnálisis rápido sueloLuis PimentelОценок пока нет
- Seminario 1 - Transporte A Travès de La MembranaДокумент42 страницыSeminario 1 - Transporte A Travès de La MembranaMILUSKA BELEN TORRES ACUÑA0% (1)
- Transportadores de glucosa: clasificación y distribuciónДокумент13 страницTransportadores de glucosa: clasificación y distribuciónPaolaPerezОценок пока нет
- Aplicaciones Medicas AcuaporinasДокумент1 страницаAplicaciones Medicas AcuaporinasEDGARDO DE JESUS BENITEZ MARTINEZОценок пока нет
- Acuaporinas: canales de agua celulares(40Документ2 страницыAcuaporinas: canales de agua celulares(40eliocesar92Оценок пока нет
- ASPECTOS DE INTERÉS SOBRE LAS ACUAPORINAS PLANTAS - 2014 - Prof Jorge A Leyva R PDFДокумент10 страницASPECTOS DE INTERÉS SOBRE LAS ACUAPORINAS PLANTAS - 2014 - Prof Jorge A Leyva R PDFPaula De Alba SurmayОценок пока нет
- Grupo 10 - Tema Difusión y Difusión A Través de Una MembranaДокумент15 страницGrupo 10 - Tema Difusión y Difusión A Través de Una MembranamjmediОценок пока нет
- NeurohipófisisДокумент36 страницNeurohipófisisIsabella Hoyos MaderaОценок пока нет
- Plantas, agua y transpiraciónДокумент68 страницPlantas, agua y transpiraciónSergio Iván López LallanaОценок пока нет
- Receptores de La VasopresinaДокумент8 страницReceptores de La VasopresinaAlexis LemaОценок пока нет
- Resumen Unidad 2 BiologiaДокумент12 страницResumen Unidad 2 BiologiaRocío PonceОценок пока нет
- Anexo 2 Formato PortadaДокумент43 страницыAnexo 2 Formato PortadaAlejandra LondoñoОценок пока нет
- Taller de TransporteДокумент2 страницыTaller de TransporteDiana Lizeth Ramirez Garcia100% (1)
- Fisiología Guyton. Capítulo 4Документ13 страницFisiología Guyton. Capítulo 4LENIN ALEXANDER BARZALLO BERREZUETAОценок пока нет
- Semana 4 - Grupo 4 - AcuaporinasДокумент15 страницSemana 4 - Grupo 4 - AcuaporinasAreli RiveraОценок пока нет
- Resistencia A La Salinidad en PlantasДокумент10 страницResistencia A La Salinidad en PlantasEduardo SosaОценок пока нет
- Biologia Molecular de Los Transport Adores de GlucosaДокумент11 страницBiologia Molecular de Los Transport Adores de GlucosaFabiola MastromatteoОценок пока нет


















































![AQUAPORINA 8- BIOQUÍMICA-24 hoy kathia[1] AVANCE HOY[1]](https://imgv2-1-f.scribdassets.com/img/document/719725637/149x198/9efd069e9c/1712164623?v=1)
![AQUAPORINA 8- BIOQUÍMICA-24 hoy kathia[1]](https://imgv2-1-f.scribdassets.com/img/document/721484460/149x198/83539ec071/1712671552?v=1)