Вам также может понравиться
- Acides nucléiques: Les Grands Articles d'UniversalisОт EverandAcides nucléiques: Les Grands Articles d'UniversalisОценок пока нет
- 1-Structure Bases, Nucléotides Et Acides Nucléiques ARN/ADNДокумент8 страниц1-Structure Bases, Nucléotides Et Acides Nucléiques ARN/ADNZineb DahmaniОценок пока нет
- 19 Thermodynamique Statique Fluides Exercices PDFДокумент1 страница19 Thermodynamique Statique Fluides Exercices PDFfsd vxxs100% (1)
- Les Effets ElectroniquesДокумент8 страницLes Effets ElectroniquesChTri Lyrics100% (1)
- TD 3Документ17 страницTD 3Hamza HamizoОценок пока нет
- Phy 5 AS23Документ98 страницPhy 5 AS23Mohamed Lemin MoustaphaОценок пока нет
- SPECTRE DE l'HYDROGENE - Modèle de BOHRДокумент4 страницыSPECTRE DE l'HYDROGENE - Modèle de BOHRoussama Labiod100% (1)
- TD 4 La ThermochimieДокумент12 страницTD 4 La ThermochimiekoumaОценок пока нет
- Impedance Comp CorrДокумент2 страницыImpedance Comp CorrmoipopОценок пока нет
- Partiel 2016 CorrigéДокумент12 страницPartiel 2016 Corrigémaurice audin100% (1)
- Laser CoursДокумент11 страницLaser CoursAhmed fattoumОценок пока нет
- TD3 Eln-2015Документ4 страницыTD3 Eln-2015hubfriОценок пока нет
- TD Chimie ThéoriqueДокумент22 страницыTD Chimie ThéoriqueKGGKFОценок пока нет
- 2.le Spectre D'atome D'hydrogeneДокумент7 страниц2.le Spectre D'atome D'hydrogeneMétatron Games100% (1)
- Résumé Chapitre 2Документ17 страницRésumé Chapitre 2Samira KaОценок пока нет
- Cours ElectricitéДокумент115 страницCours ElectricitéSannadОценок пока нет
- Cours - Chimie - Les Acides Et Les BasesДокумент6 страницCours - Chimie - Les Acides Et Les BasesDaghsni Said100% (2)
- EXERCICE 30.3-: Optique OndulatoireДокумент3 страницыEXERCICE 30.3-: Optique OndulatoireChaima HamidiОценок пока нет
- Sol TD 03Документ3 страницыSol TD 03Grine Salah EddineОценок пока нет
- FPO SMP TD Thermodynamique II 2018 2019 Serie 06 CorrectionДокумент4 страницыFPO SMP TD Thermodynamique II 2018 2019 Serie 06 CorrectionMelchisedek Meignan100% (1)
- Théorie de Champ TD 3Документ2 страницыThéorie de Champ TD 3Motive -Tech100% (1)
- Matrices Et Déterminants - TraceДокумент5 страницMatrices Et Déterminants - TraceHamza PrintoOsОценок пока нет
- Fonctions Usuelles PDFДокумент29 страницFonctions Usuelles PDFchanezОценок пока нет
- Rappel ElectrostatiqueДокумент14 страницRappel ElectrostatiqueAbdelhamid ElmadaniОценок пока нет
- Transparents Sur Les Semi-Conducteurs PDFДокумент29 страницTransparents Sur Les Semi-Conducteurs PDFKhalid GarbaОценок пока нет
- 1 2 B Enonce ExercicesДокумент8 страниц1 2 B Enonce Exerciceszineb senhajiОценок пока нет
- Solutions Fluides PDFДокумент92 страницыSolutions Fluides PDFdjemОценок пока нет
- TD3 Chimie Generale +CORRIGE 2016-2017Документ6 страницTD3 Chimie Generale +CORRIGE 2016-2017Awatif BeОценок пока нет
- Engrenages 2Документ7 страницEngrenages 2Mohamed LaliouiОценок пока нет
- Biophysique - Structure de La MatДокумент2 страницыBiophysique - Structure de La MatfghjkОценок пока нет
- Chapitre 1 Etat Cristallin PDFДокумент29 страницChapitre 1 Etat Cristallin PDFhéma tologieОценок пока нет
- Serie Avec Correction Les Spectres Atomiques Section ScientifiquesДокумент17 страницSerie Avec Correction Les Spectres Atomiques Section ScientifiquesJerbi InesОценок пока нет
- VHNPДокумент638 страницVHNPDavid BAWAОценок пока нет
- O Ch1-L'atomeДокумент2 страницыO Ch1-L'atomeGIFFARD100% (1)
- Examen 2er Semestre Mai 2017version Finale Enone Et CorrigeeДокумент8 страницExamen 2er Semestre Mai 2017version Finale Enone Et CorrigeeAziz JridiОценок пока нет
- Corrige DM5 PDFДокумент15 страницCorrige DM5 PDFBrandy OdonnellОценок пока нет
- SMPC s2 CHP 2Документ31 страницаSMPC s2 CHP 2Abdelhakim BailalОценок пока нет
- P6-5-Rayonnement D Un Dipole Oscillant PDFДокумент11 страницP6-5-Rayonnement D Un Dipole Oscillant PDFHamma MeskiniОценок пока нет
- CHAPITRE I Théorie Des ChampsДокумент15 страницCHAPITRE I Théorie Des ChampsM'pounguiОценок пока нет
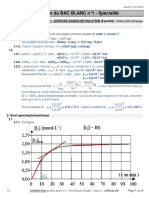
- TS - BAC Blanc N°1 Corrigé - Spécialité PDFДокумент4 страницыTS - BAC Blanc N°1 Corrigé - Spécialité PDFphytanjaОценок пока нет
- Chapitre 1 Intégration NumériqueДокумент19 страницChapitre 1 Intégration NumériqueBruno NAPOОценок пока нет
- TD Optique 3-2021Документ1 страницаTD Optique 3-2021Mohamed CHARIFОценок пока нет
- Centrale MP 2001 CorrigeДокумент10 страницCentrale MP 2001 CorrigePaolo HoguoОценок пока нет
- TD1 Chocs Corrige PHY12ab 2014Документ15 страницTD1 Chocs Corrige PHY12ab 2014AyoubIghzirОценок пока нет
- 1 Respiration 1Документ1 страница1 Respiration 1RedaОценок пока нет
- FPO SMP DS Thermodynamique I 2016-02-29 CorrectionДокумент4 страницыFPO SMP DS Thermodynamique I 2016-02-29 CorrectionHamza DickoОценок пока нет
- Corrigé Série 2Документ8 страницCorrigé Série 2Chk YahyaОценок пока нет
- Tipe Mcot 2023Документ4 страницыTipe Mcot 2023Nz 'Оценок пока нет
- A2 Niveaux D Energie AtomesДокумент11 страницA2 Niveaux D Energie AtomesfaslaОценок пока нет
- Problème OptiqueДокумент5 страницProblème OptiqueMaxwellОценок пока нет
- TD AtomДокумент6 страницTD AtomToufikJalalEddine100% (1)
- Chap 4 DiènesДокумент16 страницChap 4 DiènesWahab HoubadОценок пока нет
- PC PHYSIQUE CENTRALE 1 2002.extraitДокумент4 страницыPC PHYSIQUE CENTRALE 1 2002.extraitNadaОценок пока нет
- ProblèmesДокумент45 страницProblèmesYassine ErrakkasОценок пока нет
- Appel CNPMA'2021Документ2 страницыAppel CNPMA'2021Youcef Fermi100% (1)
- PhysiqueDéfinitions MR Haidara-1Документ8 страницPhysiqueDéfinitions MR Haidara-1Amadou TraoréОценок пока нет
- Rattrapage ST-sem3 - 2008-2009Документ5 страницRattrapage ST-sem3 - 2008-2009Rou MàissàОценок пока нет
- I - L'Adn 1. Les Constituants de l'ADNДокумент13 страницI - L'Adn 1. Les Constituants de l'ADNZakaria FerkacheОценок пока нет
- Acides NucléiquesДокумент23 страницыAcides NucléiquesKEBIR ZAHRAОценок пока нет
- Genetique1an16 AdnДокумент11 страницGenetique1an16 AdnhichamОценок пока нет
- QCM - CytogénétiqueДокумент33 страницыQCM - CytogénétiqueMouad Hilia100% (1)
- Chapitre 4-ConvertiДокумент10 страницChapitre 4-ConvertiOmaima HmamouОценок пока нет
- Chapitre 1 EleveДокумент4 страницыChapitre 1 EleveMathis LefrancОценок пока нет
- 2.bioener Metab RegulДокумент140 страниц2.bioener Metab RegulKenza KassabОценок пока нет
- TD 3 Corrigé Regulation SABRINA.2019Документ5 страницTD 3 Corrigé Regulation SABRINA.2019axelОценок пока нет
- Modulation Des Activités EnzymatiquesДокумент20 страницModulation Des Activités EnzymatiquesBMA-medecineОценок пока нет
- Méiose FR 2 PC SMДокумент1 страницаMéiose FR 2 PC SMmalakcantactproОценок пока нет
- 3 Cours Traduction PDFДокумент10 страниц3 Cours Traduction PDFMuhamedMouslimОценок пока нет
- Respiration Cellulaire PR Rajaonera TovoДокумент46 страницRespiration Cellulaire PR Rajaonera TovoFahendrenaОценок пока нет
- TD DynamiqueДокумент24 страницыTD Dynamiquesouttou sabrinaОценок пока нет
- Les-Mitochondries - Compressed 1Документ33 страницыLes-Mitochondries - Compressed 1marie dominique patricia kouassiОценок пока нет
- Metabolisme Des ProteinesДокумент32 страницыMetabolisme Des ProteinesNouaïri AmineОценок пока нет
- Exposé de Cytologie: ThèmeДокумент41 страницаExposé de Cytologie: ThèmeFaresОценок пока нет
- Cours 1 Généralités Sur La Signalisation Cellulaire 1Документ18 страницCours 1 Généralités Sur La Signalisation Cellulaire 1abdou.djat75Оценок пока нет
- 9-Le Noyau Et Le Cycle CellulaireДокумент73 страницы9-Le Noyau Et Le Cycle Cellulairemouniadahou3Оценок пока нет
- Les TélomèresДокумент1 страницаLes TélomèresgreyОценок пока нет
- Êxercic, F ': MultiplesДокумент3 страницыÊxercic, F ': MultiplessimplicitaОценок пока нет
- Chap2 PapierДокумент20 страницChap2 Papiershaddaï mavakalaОценок пока нет
- Polycopie Du Cours Régulation Des Gènes Chez Les Procaryotes M1 Génétique MM GHARZOULIДокумент64 страницыPolycopie Du Cours Régulation Des Gènes Chez Les Procaryotes M1 Génétique MM GHARZOULISarah SisiОценок пока нет
- QCM La Mitochondrie Pour Meriem1Документ5 страницQCM La Mitochondrie Pour Meriem1Kamel Ould Amrouche50% (2)
- BIO111 ChapitreIIIДокумент8 страницBIO111 ChapitreIIIBerthe Aristide Ngue NkongoОценок пока нет
- Cours Chap 5 AbsentesДокумент4 страницыCours Chap 5 AbsentesFadhel SlamaОценок пока нет
- Régulation Du Métabolisme Protéique P1Документ34 страницыRégulation Du Métabolisme Protéique P1Hicham EddaoudiОценок пока нет
- La Transcription de l'ADNДокумент4 страницыLa Transcription de l'ADNKhelОценок пока нет
- Anémies Hémolytiques Dues A Des Déficits en Enzymes Erythrocytaires Autres Que La G6PDДокумент8 страницAnémies Hémolytiques Dues A Des Déficits en Enzymes Erythrocytaires Autres Que La G6PDmanel manОценок пока нет
- L'Anatomie Et La Physiologie Pour Les Infirmier (E) SДокумент626 страницL'Anatomie Et La Physiologie Pour Les Infirmier (E) Sma meilleureversionht100% (2)
- M10574Документ90 страницM10574Mohamed Wassim M'henniОценок пока нет
- Poly Biologie MoléculaireДокумент21 страницаPoly Biologie MoléculaireRania MaddahОценок пока нет
- Sujet 2 PcemДокумент4 страницыSujet 2 PcemmedОценок пока нет
- Physio1an-Interaction Ligand Recepteur2016Документ18 страницPhysio1an-Interaction Ligand Recepteur2016AmirMohamedYozmaneОценок пока нет