Вам также может понравиться
- 5 10-12 SilbersteinДокумент54 страницы5 10-12 SilbersteinFailane LisОценок пока нет
- Tiroide e ParatiroideДокумент30 страницTiroide e ParatiroideFailane LisОценок пока нет
- Celiaquia Aep PDFДокумент10 страницCeliaquia Aep PDFGuillermo AdrianzénОценок пока нет
- 06 Elevacion IgEДокумент14 страниц06 Elevacion IgEEdgar UribeОценок пока нет
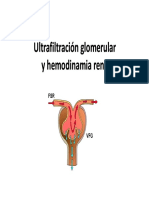
- UltrafiltradoДокумент36 страницUltrafiltradoFailane LisОценок пока нет
- ECG BasicaДокумент167 страницECG Basicaluchinmaura100% (1)
- Dialnet FitoterapiaDeLaInflamacion 4989385Документ6 страницDialnet FitoterapiaDeLaInflamacion 4989385Failane LisОценок пока нет
- Clima InstitucionalДокумент55 страницClima InstitucionaldjfulanitoОценок пока нет
- Precursores de La LogopediaДокумент7 страницPrecursores de La LogopedianoelialorenzocastilloОценок пока нет
- Acta de Intervencion Por Violacion SexualДокумент11 страницActa de Intervencion Por Violacion SexualaristacastroalexОценок пока нет
- Construccion DX de EnfermeriaДокумент6 страницConstruccion DX de EnfermeriaMartines Ami0% (1)
- Triptico de La AlcachofaДокумент3 страницыTriptico de La AlcachofaDavid NinasivinchacuelloОценок пока нет
- Distribucion Universal de MicroorganismosДокумент37 страницDistribucion Universal de MicroorganismosLuis Aarón Cabello Candela0% (1)
- Prevención de Riesgos ProfesionalesДокумент6 страницPrevención de Riesgos ProfesionalesArenas Ruiz KarinaОценок пока нет
- Aupr - Tema 3Документ16 страницAupr - Tema 3Jesús Del Río RodríguezОценок пока нет
- Factores Que Afectan A La SaludДокумент4 страницыFactores Que Afectan A La SaludEderth CorderoОценок пока нет
- Hiperreactividad BronquialДокумент3 страницыHiperreactividad BronquialAlejandro Seneca HernandezОценок пока нет
- Local de Elaboracion de Alimentos Con Consumo Al PasoДокумент3 страницыLocal de Elaboracion de Alimentos Con Consumo Al PasoKenyiro Angelo Asada SolariОценок пока нет
- BassedasДокумент5 страницBassedasMarcela QuarchioniОценок пока нет
- Consecuencias de La ConquistaДокумент2 страницыConsecuencias de La ConquistabetbetbetОценок пока нет
- Sílabo Corregido Relaciones HumanasДокумент5 страницSílabo Corregido Relaciones HumanasRomeen Aguirre ZambranoОценок пока нет
- Neurocisticercosis en Cajamarca (Dr. Juan Salazar Pajares) 1Документ16 страницNeurocisticercosis en Cajamarca (Dr. Juan Salazar Pajares) 1Juan C. Salazar PajaresОценок пока нет
- Campos de Intervencion Del Trabajo SocialДокумент4 страницыCampos de Intervencion Del Trabajo SocialAxa Capital100% (1)
- Tarea Individual Radiologia 2019Документ20 страницTarea Individual Radiologia 2019Diana CarranzaОценок пока нет
- Declaración Jurada de SaludДокумент1 страницаDeclaración Jurada de SaludMarco Antonio Porras PajueloОценок пока нет
- Carta-Descriptiva-Ec0217-Ec0301-Producto Nom-019-STPS-2011Документ11 страницCarta-Descriptiva-Ec0217-Ec0301-Producto Nom-019-STPS-2011Jdionicio. Surestegmail.com DionicioОценок пока нет
- Sesión Aprendiendo A Reconocer Situaciones de Riesgo Dimensión PersonalДокумент3 страницыSesión Aprendiendo A Reconocer Situaciones de Riesgo Dimensión PersonalJose Luis Quispe FernandezОценок пока нет
- Guia 1 Corazon y PulmonДокумент4 страницыGuia 1 Corazon y PulmonNataly Labraña CamposОценок пока нет
- Proyecto Final PHPДокумент2 страницыProyecto Final PHPDiego MolinaОценок пока нет
- Reanimación Cardiopulmonar RCP EstudiantesДокумент18 страницReanimación Cardiopulmonar RCP EstudiantesAndrea DuránОценок пока нет
- Capitulo 7Документ14 страницCapitulo 7Melani MiglioriniОценок пока нет
- 3.indagamos Sobre La Presencia de Hierro en Los AlimentosДокумент7 страниц3.indagamos Sobre La Presencia de Hierro en Los AlimentosMaría Isidora Aquino SosaОценок пока нет
- Grupo 8 - Guion - Me Doy Una OportunidadДокумент10 страницGrupo 8 - Guion - Me Doy Una OportunidadRayza Andrea Callupe PhoccoОценок пока нет
- Ficha Descriptiva de Silla de Ruedas NeurológicaДокумент8 страницFicha Descriptiva de Silla de Ruedas NeurológicaDeathV4Оценок пока нет
- Malformaciones Congenitas NeurologicasДокумент9 страницMalformaciones Congenitas NeurologicasStephany Antonela Monrroy SisnerosОценок пока нет
- Ejercicios de GenealogíaДокумент10 страницEjercicios de GenealogíaKatherine Paredes GomezОценок пока нет
- ExamenFinal Psicopatologia Del Infante 2Документ26 страницExamenFinal Psicopatologia Del Infante 2Esteban SanchezОценок пока нет