Вам также может понравиться
- Microbial Proteomics: Development in Technologies and ApplicationsОт EverandMicrobial Proteomics: Development in Technologies and ApplicationsОценок пока нет
- 2012-Bti ABC Transporter Toxin ModeДокумент8 страниц2012-Bti ABC Transporter Toxin ModeDwiОценок пока нет
- The Role of Products of the Histocompatibility Gene Complex in Immune ResponsesОт EverandThe Role of Products of the Histocompatibility Gene Complex in Immune ResponsesОценок пока нет
- Antibiotic Resistance Mechanisms in Bacteria: Biochemical and Genetic AspectsДокумент11 страницAntibiotic Resistance Mechanisms in Bacteria: Biochemical and Genetic AspectsD Wisam NajmОценок пока нет
- Ijms 20 04007Документ16 страницIjms 20 04007Friska SiamiОценок пока нет
- Insights Into The Mode of Action of Chitosan As An Antibacterial CompoundДокумент10 страницInsights Into The Mode of Action of Chitosan As An Antibacterial CompoundsthiemchiprodeОценок пока нет
- 2-IJMS 2020-7521 PeptaibolsДокумент18 страниц2-IJMS 2020-7521 PeptaibolsImen GuerroudjОценок пока нет
- Antioxidants 11 02186Документ12 страницAntioxidants 11 02186Igor BaltaОценок пока нет
- Johansson, 2011Документ18 страницJohansson, 2011Cáceres EmiОценок пока нет
- 10 1016@j Cellsig 2020 109867Документ34 страницы10 1016@j Cellsig 2020 109867VivianKlembergОценок пока нет
- Silver Nanoparticles As Potential Antibacterial AgentsДокумент20 страницSilver Nanoparticles As Potential Antibacterial AgentsFENFOGОценок пока нет
- Systems Biology Approaches To Bioremediation: Vı Ctor de LorenzoДокумент11 страницSystems Biology Approaches To Bioremediation: Vı Ctor de LorenzoCristian Saldaña CabreraОценок пока нет
- 2020 - Bacterial Lipase Triggers The Release of Antibiotics From Digestible LiquidДокумент15 страниц2020 - Bacterial Lipase Triggers The Release of Antibiotics From Digestible LiquidHe ZeeОценок пока нет
- Azitromicina DocumentoДокумент12 страницAzitromicina DocumentoAlex Enrique Acosta RamosОценок пока нет
- Iriti 2008Документ6 страницIriti 2008Gustavo HoppeОценок пока нет
- An Optimized Anti-Adherence and Anti-Biofilm Assay Case Study of Zinc Oxide Nanoparticles Versus MRSA BiofilmДокумент6 страницAn Optimized Anti-Adherence and Anti-Biofilm Assay Case Study of Zinc Oxide Nanoparticles Versus MRSA BiofilmKareem IsmailОценок пока нет
- Introduction and Overview Antibiotics and Chemotherapeutic Agents Micro 260 Chapter 12 NotesДокумент8 страницIntroduction and Overview Antibiotics and Chemotherapeutic Agents Micro 260 Chapter 12 NotesNEHA DIXITОценок пока нет
- Chitosan Antimicrobial and Eliciting Properties For Pest ControlДокумент20 страницChitosan Antimicrobial and Eliciting Properties For Pest ControlTesfabirhan MiracleОценок пока нет
- ART Action and Resistance Mechanisms of Antibiotics A Guide For CliniciansДокумент16 страницART Action and Resistance Mechanisms of Antibiotics A Guide For CliniciansHECTORIBZAN ACERO SANDOVALОценок пока нет
- Pengawet KitosanДокумент15 страницPengawet KitosanCindy AnggraeniОценок пока нет
- I - Inhibition of Streptococcus Mutans Biofilms With Bacterial-Derived Outer Membrane VesiclesДокумент12 страницI - Inhibition of Streptococcus Mutans Biofilms With Bacterial-Derived Outer Membrane VesiclesHime IbaraОценок пока нет
- Compounds Kolkisin 3Документ11 страницCompounds Kolkisin 3Evi WulansariОценок пока нет
- Microbiome and Diseases: Pathogen Infection: Christine Josenhans and Guntram A. GrasslДокумент22 страницыMicrobiome and Diseases: Pathogen Infection: Christine Josenhans and Guntram A. GrasslÂngela Souza BarroqueiroОценок пока нет
- Printed in Great Britain. All Rights Reserved, ©: Biotech. Adv. Vol. 9, Pp. 217-240,1991Документ24 страницыPrinted in Great Britain. All Rights Reserved, ©: Biotech. Adv. Vol. 9, Pp. 217-240,1991sebastian floresОценок пока нет
- Tema 3 - Inmune Responses To ImplantsДокумент18 страницTema 3 - Inmune Responses To ImplantsIsrael GonzálezОценок пока нет
- A General Model For Toxin-Antitoxin Module Dynamics Can Explain Persister Cell Formation in E. ColiДокумент17 страницA General Model For Toxin-Antitoxin Module Dynamics Can Explain Persister Cell Formation in E. ColiДушан МарковићОценок пока нет
- Chemotaxis Review EthДокумент23 страницыChemotaxis Review EthPracren TamneОценок пока нет
- Mechanisms of Antibiotic ActionДокумент15 страницMechanisms of Antibiotic ActionMarc Imhotep Cray, M.D.Оценок пока нет
- Food Applications of Chitin and ChitosansДокумент15 страницFood Applications of Chitin and ChitosansRaul Diaz TorresОценок пока нет
- Antibiotics: Classification Framework and Chemical Biology of Tetracycline-Structure-Based DrugsДокумент13 страницAntibiotics: Classification Framework and Chemical Biology of Tetracycline-Structure-Based DrugsMuthia FadhilaОценок пока нет
- Microbial Virulence Factors: Molecular SciencesДокумент6 страницMicrobial Virulence Factors: Molecular SciencesAnasОценок пока нет
- Probiotic Activities of Lcr35 in Vitro Adherence To Intestinal Cells and Antimicrobial PropertiesДокумент7 страницProbiotic Activities of Lcr35 in Vitro Adherence To Intestinal Cells and Antimicrobial PropertiesMaria TejedaОценок пока нет
- Review of Biofilm Inhibiting PhytochemicalsДокумент26 страницReview of Biofilm Inhibiting PhytochemicalsMichal SladekОценок пока нет
- Riddle of Biofilm Resistance by Kim LewisДокумент9 страницRiddle of Biofilm Resistance by Kim LewisribozymesОценок пока нет
- Toxins: BT Toxin Modification For Enhanced EfficacyДокумент23 страницыToxins: BT Toxin Modification For Enhanced EfficacyCamila VelandiaОценок пока нет
- Chemik 8 2012 02Документ5 страницChemik 8 2012 02mahmoud shirbiniОценок пока нет
- Synbiotic Combination of Lactobacillus Rhamnosus NCDC 298 and Short Chain Fructooligosaccharides Prevents Enterotoxigenic Escherichia Coli InfectionДокумент6 страницSynbiotic Combination of Lactobacillus Rhamnosus NCDC 298 and Short Chain Fructooligosaccharides Prevents Enterotoxigenic Escherichia Coli InfectionDidier Alexis Rosales MosqueraОценок пока нет
- Antibiotics 01 00001Документ13 страницAntibiotics 01 00001Ravi KantОценок пока нет
- Optimization of Cultural Conditions For The Production of Antifungal Chitinase by Streptomyces SporovirgulisДокумент7 страницOptimization of Cultural Conditions For The Production of Antifungal Chitinase by Streptomyces SporovirgulisDella AzzuraОценок пока нет
- Accepted Manuscript: Carbohydrate PolymersДокумент28 страницAccepted Manuscript: Carbohydrate PolymersJumardinОценок пока нет
- The Immune Escape Mechanisms of Mycobacterium Tuberculosis: Molecular SciencesДокумент18 страницThe Immune Escape Mechanisms of Mycobacterium Tuberculosis: Molecular SciencesMaria Gabriella Ziglio da SilvaОценок пока нет
- HW 0440Документ8 страницHW 0440JaanvhiОценок пока нет
- Gut Microbiome Modulates Efficacy of Immune Checkpoint InhibitorsДокумент10 страницGut Microbiome Modulates Efficacy of Immune Checkpoint InhibitorsGusti FerriОценок пока нет
- Molecules: Antibiotic Discovery: Combatting Bacterial Resistance in Cells and in Biofilm CommunitiesДокумент13 страницMolecules: Antibiotic Discovery: Combatting Bacterial Resistance in Cells and in Biofilm CommunitiesluluksОценок пока нет
- Lichen Secondary Metabolites As Potential Antibiotic Agents: 3.1 Needing For New Antibiotics From NatureДокумент24 страницыLichen Secondary Metabolites As Potential Antibiotic Agents: 3.1 Needing For New Antibiotics From NatureIrfaan SalzabilОценок пока нет
- 7 Exploiting Plant Innate Immunity To Protect Crops Against Biotic Stress: Chitosaccharides As Natural and Suitable Candidates For This PurposeДокумент29 страниц7 Exploiting Plant Innate Immunity To Protect Crops Against Biotic Stress: Chitosaccharides As Natural and Suitable Candidates For This PurposeJose Miguel Soto HerediaОценок пока нет
- Pared Celular M. TuberculosisДокумент13 страницPared Celular M. TuberculosisJose Maria RojasОценок пока нет
- Omics Breakthroughs For Environmental MicrobiologyДокумент16 страницOmics Breakthroughs For Environmental Microbiologyabcder1234Оценок пока нет
- Bacteriocins Modes of Action and Potentials in Food Preservation and Control of Food Poisoning 1995 International Journal of Food MicrobiologyДокумент17 страницBacteriocins Modes of Action and Potentials in Food Preservation and Control of Food Poisoning 1995 International Journal of Food MicrobiologyNissa MarlindaОценок пока нет
- Perz-Diaz2015, Silver Nanoparticles With Antimicrobial Activities Against Streptococcus Mutans and Their Cytotoxic EffectДокумент7 страницPerz-Diaz2015, Silver Nanoparticles With Antimicrobial Activities Against Streptococcus Mutans and Their Cytotoxic EffectPhuong ThaoОценок пока нет
- Fimmu 06 00555Документ20 страницFimmu 06 00555www.henry1020Оценок пока нет
- Elimnation of MycotixinsДокумент11 страницElimnation of MycotixinsSrivatsava RajagopalanОценок пока нет
- Inadequate Timing Between Corticosteroid and Antibiotics Applications Increases Mortality Due To SepsisДокумент7 страницInadequate Timing Between Corticosteroid and Antibiotics Applications Increases Mortality Due To SepsisDwi afrianti Rahma putriОценок пока нет
- An in Vitro Cytotoxicity of A Novel PH Sensitive Lecti - 2021 - Journal of DrugДокумент14 страницAn in Vitro Cytotoxicity of A Novel PH Sensitive Lecti - 2021 - Journal of DrugRiki WijayaОценок пока нет
- Biomedical and Therapeutic Applications of Biosurfactants: Chapter #Документ14 страницBiomedical and Therapeutic Applications of Biosurfactants: Chapter #Trainer NIAОценок пока нет
- Mammalian Synthetic Biology - From Tools To Therapies: Dominique Aubel, and Martin FusseneggerДокумент14 страницMammalian Synthetic Biology - From Tools To Therapies: Dominique Aubel, and Martin FusseneggerChickencito PerezОценок пока нет
- Cells: Perspectives For Glyco-Engineering of Recombinant Biopharmaceuticals From MicroalgaeДокумент26 страницCells: Perspectives For Glyco-Engineering of Recombinant Biopharmaceuticals From MicroalgaeVaibhav LonganiОценок пока нет
- LeptocarpinaДокумент7 страницLeptocarpinaJ Nicolas MJОценок пока нет
- Chhatwal 2002Документ4 страницыChhatwal 2002Rin ChanОценок пока нет
- Polymers: Biomedical Applications of Chitosan and Its Derivative NanoparticlesДокумент17 страницPolymers: Biomedical Applications of Chitosan and Its Derivative Nanoparticlesfrancisco cadenaОценок пока нет
- MITOCHONDRONДокумент3 страницыMITOCHONDRONPhoebe Magdurulan100% (1)
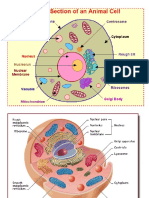
- CellsДокумент24 страницыCellsapi-95529788100% (1)
- Functional Anatomy of The Prokaryotic CellДокумент22 страницыFunctional Anatomy of The Prokaryotic Cellm umair zahirОценок пока нет
- Lesson-3-Prokaryotes-and-Eukaryotes CellsДокумент14 страницLesson-3-Prokaryotes-and-Eukaryotes CellsKivo ZoshikoroОценок пока нет
- Cell Size and ShapeДокумент23 страницыCell Size and ShapenmsarmaОценок пока нет
- The Parts of The Cell and Their FunctionsДокумент2 страницыThe Parts of The Cell and Their Functionsraphael100% (1)
- Csir - Ugc Net Syllabus - Life Sciences (LS) : 1. Molecules and Their Interaction Relavent To BiologyДокумент8 страницCsir - Ugc Net Syllabus - Life Sciences (LS) : 1. Molecules and Their Interaction Relavent To BiologyEr Purushottam PalОценок пока нет
- Extra Questions For Class 9th - CH 5 The Fundamental Unit of Life Science Study Rankers PDFДокумент4 страницыExtra Questions For Class 9th - CH 5 The Fundamental Unit of Life Science Study Rankers PDFsssОценок пока нет
- Question Bank of Biochemistry PDFДокумент214 страницQuestion Bank of Biochemistry PDFwaadОценок пока нет
- Why Are Cells So Small MyДокумент2 страницыWhy Are Cells So Small Myapi-521781723Оценок пока нет
- The Effect of Alcohols On Biological MembranesДокумент6 страницThe Effect of Alcohols On Biological Membranesangel_1_Оценок пока нет
- Unit 3 Packet Carolyn StudentДокумент28 страницUnit 3 Packet Carolyn Studentmrsblennau100% (2)
- ANTIOXIDANT MEXIDOL-The Main Neuropsychotropic EffectДокумент21 страницаANTIOXIDANT MEXIDOL-The Main Neuropsychotropic Effectmatsapo100% (1)
- PHYSIOLOGY Lecture Notes Chapter 1Документ9 страницPHYSIOLOGY Lecture Notes Chapter 1Aubrey Unique EvangelistaОценок пока нет
- Anaphy Pre LimДокумент13 страницAnaphy Pre LimKyle M. BayangosОценок пока нет
- Penetrate BarriersДокумент6 страницPenetrate BarriersCatherine TangОценок пока нет
- General Biology 1 Prokaryotic Vs Eukaryotic Cells Quarter 1, Module 2aДокумент10 страницGeneral Biology 1 Prokaryotic Vs Eukaryotic Cells Quarter 1, Module 2aSam KimОценок пока нет
- Antiseptics and Disinfectants ProductsДокумент25 страницAntiseptics and Disinfectants ProductsAngela Tamayo100% (1)
- PracticalsДокумент33 страницыPracticalsNo NameОценок пока нет
- Cell - Structure and Function Class 8 Notes: CellsДокумент26 страницCell - Structure and Function Class 8 Notes: CellsPrabal SinghОценок пока нет
- Cell Defense Worksheet OnlineДокумент3 страницыCell Defense Worksheet OnlineFrancisco Solis AilonОценок пока нет
- Membrane Structure and Function PresentationДокумент13 страницMembrane Structure and Function PresentationUnus AnnusОценок пока нет
- Diffusion Osmosis and Active TransportДокумент18 страницDiffusion Osmosis and Active TransportThompson Ta Ta JevonОценок пока нет
- International Journal of Scientific and Innovative Research 2015 3 (1) P-ISSN 2347-2189, E - ISSN 2347-4971Документ172 страницыInternational Journal of Scientific and Innovative Research 2015 3 (1) P-ISSN 2347-2189, E - ISSN 2347-4971Dr.B.R.PandeyОценок пока нет
- CH4306 Lec 00 Biologics ProductionДокумент66 страницCH4306 Lec 00 Biologics ProductionstarlcОценок пока нет
- General Biology 1: First Semester-Quarter 1Документ26 страницGeneral Biology 1: First Semester-Quarter 1sjhdjshakjОценок пока нет
- Membrane-Transport WebquestДокумент2 страницыMembrane-Transport WebquestCourtlynn RodgersОценок пока нет
- How Herbicides Work - Biology To Application - University of AlbertaДокумент148 страницHow Herbicides Work - Biology To Application - University of AlbertaJuliano LorenzettiОценок пока нет
- Bacteria OutlineДокумент4 страницыBacteria OutlineCinds SumintanОценок пока нет
- Problems LipidsДокумент2 страницыProblems LipidsDittha Rieztya Phyramid0% (1)
- Hero Found: The Greatest POW Escape of the Vietnam WarОт EverandHero Found: The Greatest POW Escape of the Vietnam WarРейтинг: 4 из 5 звезд4/5 (19)
- Dirt to Soil: One Family’s Journey into Regenerative AgricultureОт EverandDirt to Soil: One Family’s Journey into Regenerative AgricultureРейтинг: 5 из 5 звезд5/5 (125)
- The End of Craving: Recovering the Lost Wisdom of Eating WellОт EverandThe End of Craving: Recovering the Lost Wisdom of Eating WellРейтинг: 4.5 из 5 звезд4.5/5 (82)
- Sully: The Untold Story Behind the Miracle on the HudsonОт EverandSully: The Untold Story Behind the Miracle on the HudsonРейтинг: 4 из 5 звезд4/5 (103)
- The Fabric of Civilization: How Textiles Made the WorldОт EverandThe Fabric of Civilization: How Textiles Made the WorldРейтинг: 4.5 из 5 звезд4.5/5 (58)
- Faster: How a Jewish Driver, an American Heiress, and a Legendary Car Beat Hitler's BestОт EverandFaster: How a Jewish Driver, an American Heiress, and a Legendary Car Beat Hitler's BestРейтинг: 4 из 5 звезд4/5 (28)
- The Future of Geography: How the Competition in Space Will Change Our WorldОт EverandThe Future of Geography: How the Competition in Space Will Change Our WorldРейтинг: 4 из 5 звезд4/5 (6)
- When the Heavens Went on Sale: The Misfits and Geniuses Racing to Put Space Within ReachОт EverandWhen the Heavens Went on Sale: The Misfits and Geniuses Racing to Put Space Within ReachРейтинг: 3.5 из 5 звезд3.5/5 (6)
- The Beekeeper's Lament: How One Man and Half a Billion Honey Bees Help Feed AmericaОт EverandThe Beekeeper's Lament: How One Man and Half a Billion Honey Bees Help Feed AmericaОценок пока нет
- ChatGPT Money Machine 2024 - The Ultimate Chatbot Cheat Sheet to Go From Clueless Noob to Prompt Prodigy Fast! Complete AI Beginner’s Course to Catch the GPT Gold Rush Before It Leaves You BehindОт EverandChatGPT Money Machine 2024 - The Ultimate Chatbot Cheat Sheet to Go From Clueless Noob to Prompt Prodigy Fast! Complete AI Beginner’s Course to Catch the GPT Gold Rush Before It Leaves You BehindОценок пока нет
- The Intel Trinity: How Robert Noyce, Gordon Moore, and Andy Grove Built the World's Most Important CompanyОт EverandThe Intel Trinity: How Robert Noyce, Gordon Moore, and Andy Grove Built the World's Most Important CompanyОценок пока нет
- The Technology Trap: Capital, Labor, and Power in the Age of AutomationОт EverandThe Technology Trap: Capital, Labor, and Power in the Age of AutomationРейтинг: 4.5 из 5 звезд4.5/5 (46)
- Pale Blue Dot: A Vision of the Human Future in SpaceОт EverandPale Blue Dot: A Vision of the Human Future in SpaceРейтинг: 4.5 из 5 звезд4.5/5 (588)
- Four Battlegrounds: Power in the Age of Artificial IntelligenceОт EverandFour Battlegrounds: Power in the Age of Artificial IntelligenceРейтинг: 5 из 5 звезд5/5 (5)
- Permaculture for the Rest of Us: Abundant Living on Less than an AcreОт EverandPermaculture for the Rest of Us: Abundant Living on Less than an AcreРейтинг: 4.5 из 5 звезд4.5/5 (33)
- The Book of the Moon: A Guide to Our Closest NeighborОт EverandThe Book of the Moon: A Guide to Our Closest NeighborРейтинг: 4.5 из 5 звезд4.5/5 (11)
- Restoration Agriculture: Real-World Permaculture for FarmersОт EverandRestoration Agriculture: Real-World Permaculture for FarmersРейтинг: 4.5 из 5 звезд4.5/5 (86)
- Project Management All-in-One For DummiesОт EverandProject Management All-in-One For DummiesРейтинг: 5 из 5 звезд5/5 (6)
- Reality+: Virtual Worlds and the Problems of PhilosophyОт EverandReality+: Virtual Worlds and the Problems of PhilosophyРейтинг: 4 из 5 звезд4/5 (24)
- Process Plant Equipment: Operation, Control, and ReliabilityОт EverandProcess Plant Equipment: Operation, Control, and ReliabilityРейтинг: 5 из 5 звезд5/5 (1)