Вам также может понравиться
- Atlas of Early Zebrafish Brain Development: A Tool for Molecular NeurogeneticsОт EverandAtlas of Early Zebrafish Brain Development: A Tool for Molecular NeurogeneticsОценок пока нет
- (Neuromethods 50) Kevin L. Brown, Diana S. Woodruff-Pak (Auth.), Jacob Raber (Eds.) - Animal Models of Behavioral Analysis-Humana Press (2011)Документ359 страниц(Neuromethods 50) Kevin L. Brown, Diana S. Woodruff-Pak (Auth.), Jacob Raber (Eds.) - Animal Models of Behavioral Analysis-Humana Press (2011)Héctor Enrique Hernández Hinojiante100% (1)
- Electroencephalography: Electroencephalography-based mind reading is no longer just a fantasy; it may be a reality in the near futureОт EverandElectroencephalography: Electroencephalography-based mind reading is no longer just a fantasy; it may be a reality in the near futureОценок пока нет
- Molecular Biology of Human Brain Tumors: November 2017Документ39 страницMolecular Biology of Human Brain Tumors: November 2017Aleksandar Dimovski100% (1)
- Techniques in Neuroanatomical ResearchДокумент404 страницыTechniques in Neuroanatomical ResearchVlad TomaОценок пока нет
- VNG 2013Документ6 страницVNG 2013AmineОценок пока нет
- Biological Bases of Behaviour.: Lecture 7: Techniques For Understanding Brain Structure & FunctionДокумент22 страницыBiological Bases of Behaviour.: Lecture 7: Techniques For Understanding Brain Structure & FunctionVijendraAgarОценок пока нет
- Clinical Electroencephalography and Topographic Brain Mapping - Technology and Practice (PDFDrive)Документ313 страницClinical Electroencephalography and Topographic Brain Mapping - Technology and Practice (PDFDrive)marceloheitor100% (1)
- The New Clasification ILAE 2017Документ6 страницThe New Clasification ILAE 2017Ami D ALОценок пока нет
- ILAE Classification of Seizures and EpilepsyДокумент19 страницILAE Classification of Seizures and EpilepsyIRENA GENI100% (1)
- Neuroimaging Biomarkers in Alzheimer's DiseaseДокумент136 страницNeuroimaging Biomarkers in Alzheimer's DiseaseInternational Medical PublisherОценок пока нет
- Neuroglia: Non-Neuronal Cells of The Nervous SystemДокумент18 страницNeuroglia: Non-Neuronal Cells of The Nervous SystemPro fatherОценок пока нет
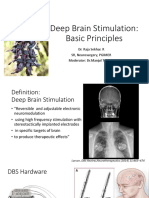
- Deep Brain Stimulation: Basic Principles: Dr. Raja Sekhar. R SR, Neurosurgery, PGIMER Moderator: DR - Manjul TripathiДокумент46 страницDeep Brain Stimulation: Basic Principles: Dr. Raja Sekhar. R SR, Neurosurgery, PGIMER Moderator: DR - Manjul Tripathijesus20100Оценок пока нет
- 18 - Thalamus and Limbic System (Edited)Документ22 страницы18 - Thalamus and Limbic System (Edited)Fotocopias LulisОценок пока нет
- Reconstruction of Virtual Neural Circuits in An Insect BrainДокумент8 страницReconstruction of Virtual Neural Circuits in An Insect BrainnithiananthiОценок пока нет
- Neurohormonal Regulation of Body FunctionsДокумент34 страницыNeurohormonal Regulation of Body FunctionsjimmyОценок пока нет
- Neuro ImagingДокумент48 страницNeuro ImagingSeiska MegaОценок пока нет
- (Natoc 1Документ495 страниц(Natoc 1Entre100% (1)
- Book Matter - AtlasOf EpilepsiesДокумент55 страницBook Matter - AtlasOf EpilepsiesDhamara AdhityaОценок пока нет
- A. Orlando Ortiz (Auth.) - Image-Guided Percutaneous Spine Biopsy-Springer International Publishing (2017)Документ290 страницA. Orlando Ortiz (Auth.) - Image-Guided Percutaneous Spine Biopsy-Springer International Publishing (2017)RUCHIR UTTAMОценок пока нет
- (Springer Series in Computational Neuroscience 3) Michael Okun, Alik Mokeichev, Yonatan Katz (Auth.), Kre¿Imir Josic, Jonathan Rubin, Manuel Matias, Ranulfo Romo (Eds.)-Coherent Behavior in Neuronal NДокумент311 страниц(Springer Series in Computational Neuroscience 3) Michael Okun, Alik Mokeichev, Yonatan Katz (Auth.), Kre¿Imir Josic, Jonathan Rubin, Manuel Matias, Ranulfo Romo (Eds.)-Coherent Behavior in Neuronal Nlig0% (1)
- B5W1L9.Peripheral Neuropathy - Lecture Notes 12Документ4 страницыB5W1L9.Peripheral Neuropathy - Lecture Notes 12mihalcea alinОценок пока нет
- Frontal Lobe Syndromes: BackgroundДокумент2 страницыFrontal Lobe Syndromes: BackgroundFausiah Ulva MОценок пока нет
- Brain Facts: A Primer On The Brain and Nervous SystemДокумент64 страницыBrain Facts: A Primer On The Brain and Nervous SystemZakir UllahОценок пока нет
- Normal Brain Anatomy On Magnetic Resonance ImagingДокумент9 страницNormal Brain Anatomy On Magnetic Resonance ImagingAnonymous vnv6QFОценок пока нет
- Advances in Electrodermal Activity Processing With Applications For Mental HealthДокумент148 страницAdvances in Electrodermal Activity Processing With Applications For Mental HealthMahdi HosseiniОценок пока нет
- Basic Brain Cross Sectional and Surface Anatomy and Function LocalizationДокумент184 страницыBasic Brain Cross Sectional and Surface Anatomy and Function Localizationmandeep axonОценок пока нет
- Carrillo Mora PDFДокумент10 страницCarrillo Mora PDFLukas GonzalezОценок пока нет
- Reinhard L. Friede M.D. (Auth.) - Developmental Neuropathology-Springer Vienna (1975) PDFДокумент535 страницReinhard L. Friede M.D. (Auth.) - Developmental Neuropathology-Springer Vienna (1975) PDFMuhammad HabiburrahmanОценок пока нет
- 19-Microendoscopic Lumbar DiscectomyДокумент8 страниц19-Microendoscopic Lumbar DiscectomyNewton IssacОценок пока нет
- Thoracic Endoscopic Spine Surgery A ComprehensiveДокумент11 страницThoracic Endoscopic Spine Surgery A Comprehensivesanjay chhawraОценок пока нет
- BPPVДокумент10 страницBPPVRizki MaulanaОценок пока нет
- Review of Anatomy of The EarДокумент16 страницReview of Anatomy of The EarSahrish IqbalОценок пока нет
- The Cerebellum-8 PDFДокумент83 страницыThe Cerebellum-8 PDFlovelots1234Оценок пока нет
- DBS2008Документ48 страницDBS2008Carol Artigas Gómez100% (1)
- CerebrumДокумент59 страницCerebrumKamlesh DuggaОценок пока нет
- MEG - An Introduction To MethodsДокумент449 страницMEG - An Introduction To MethodsSayako87Оценок пока нет
- Vant Hof SpringerProtocols 2019 V2finalДокумент38 страницVant Hof SpringerProtocols 2019 V2finalSusi SusantiОценок пока нет
- 2016 Annual Report: Multiscale Approaches To Move Science ForwardДокумент16 страниц2016 Annual Report: Multiscale Approaches To Move Science ForwardalleninstituteОценок пока нет
- #Human BrainДокумент28 страниц#Human BrainDharaa GuptaОценок пока нет
- Intracerebral Hemorrhage TherapeuticsДокумент12 страницIntracerebral Hemorrhage TherapeuticsPedro Jerry Sevilla RomeroОценок пока нет
- Neuro AnatomyДокумент26 страницNeuro Anatomymeryclave345Оценок пока нет
- Self Cingulo PosteriorДокумент7 страницSelf Cingulo PosteriorJose ZapataОценок пока нет
- Seizures, Headache, Itis Etc Leah KellyДокумент16 страницSeizures, Headache, Itis Etc Leah Kellyathena19Оценок пока нет
- SIR Model Excel2Документ8 страницSIR Model Excel2Eric ChanОценок пока нет
- CerebellumДокумент51 страницаCerebellumLiya ThaaraОценок пока нет
- Snell Reticular Formation and Limbic SystemДокумент9 страницSnell Reticular Formation and Limbic SystemFrnz RiveraОценок пока нет
- The Second Edition Expert Consensus Statements On The Use of IVIg in NeurologyДокумент134 страницыThe Second Edition Expert Consensus Statements On The Use of IVIg in NeurologyRonald DewaОценок пока нет
- Overview and Assessment of Balance Functions: Electronystagmography/ VideonystagmographyДокумент48 страницOverview and Assessment of Balance Functions: Electronystagmography/ VideonystagmographyArun KumarОценок пока нет
- Animal Models in Disease Research: Honours Department of Pathology University of Melbourne Theo MantamadiotisДокумент35 страницAnimal Models in Disease Research: Honours Department of Pathology University of Melbourne Theo MantamadiotisRiv sashОценок пока нет
- Basal Ganglia - An Integrative View PDFДокумент124 страницыBasal Ganglia - An Integrative View PDFnaresh sharma100% (1)
- Noninvasive Human Brain Stimulation: Timothy Wagner, Antoni Valero-Cabre, and Alvaro Pascual-LeoneДокумент41 страницаNoninvasive Human Brain Stimulation: Timothy Wagner, Antoni Valero-Cabre, and Alvaro Pascual-LeoneGhinea IulianaОценок пока нет
- Power Point Presentation SKELETAL SYSTEMДокумент75 страницPower Point Presentation SKELETAL SYSTEMaprillianlozanoОценок пока нет
- 01-Wk1 - (05 - 10 - 06) LECTURE 1 - STRUCTURE AND FUNCTION OF THE BRAIN-1160379353Документ26 страниц01-Wk1 - (05 - 10 - 06) LECTURE 1 - STRUCTURE AND FUNCTION OF THE BRAIN-1160379353Red Fool100% (1)
- Kandel Et Al. 2014 The Molecular and Systems Biology of MemoryДокумент24 страницыKandel Et Al. 2014 The Molecular and Systems Biology of MemoryJava UDОценок пока нет
- Percutaneous Tracheostomy in Critically Patients: Giuseppe Servillo Paolo PelosiДокумент162 страницыPercutaneous Tracheostomy in Critically Patients: Giuseppe Servillo Paolo PelosiRafael Bagus100% (1)
- Femto TechnologyДокумент11 страницFemto TechnologySanthosh Salem100% (1)
- Neural DarwinismДокумент11 страницNeural DarwinismRodrigoPristaОценок пока нет
- Cerebrospinal FluidДокумент13 страницCerebrospinal Fluidsakshi panwarОценок пока нет
- Neuroimmunology - 1Документ41 страницаNeuroimmunology - 1Michael Mazvidza100% (1)
- MF - 2-2022-Ferenc Et AlДокумент13 страницMF - 2-2022-Ferenc Et AlSinisa RisticОценок пока нет
- UZ Donje Suplje Vene U Procjeni Volumena Cirkulisuce KrviДокумент27 страницUZ Donje Suplje Vene U Procjeni Volumena Cirkulisuce KrviSinisa RisticОценок пока нет
- Marinedrugs 20 00397 v2Документ51 страницаMarinedrugs 20 00397 v2Sinisa RisticОценок пока нет
- Interpreting ECGs A Practical Approach 1st EdДокумент769 страницInterpreting ECGs A Practical Approach 1st EdArtem Trunov100% (2)
- Fphy 08 00047Документ29 страницFphy 08 00047Sinisa RisticОценок пока нет
- Pocus Point of Care Ultrazvuk Bedside UltrazvukДокумент23 страницыPocus Point of Care Ultrazvuk Bedside UltrazvukSinisa RisticОценок пока нет
- Synapses, Neurons and Brains - CourseraДокумент6 страницSynapses, Neurons and Brains - CourseraSinisa Ristic0% (1)
- 1107-1120 - Seminar On Big Data Application Impact and Telecom Business AnalysisДокумент3 страницы1107-1120 - Seminar On Big Data Application Impact and Telecom Business AnalysisSinisa RisticОценок пока нет
- Nonverbal CommunicationДокумент5 страницNonverbal CommunicationkalimeriОценок пока нет
- 1107-1120 - Seminar On Big Data Application Impact and Telecom Business AnalysisДокумент3 страницы1107-1120 - Seminar On Big Data Application Impact and Telecom Business AnalysisSinisa RisticОценок пока нет
- Advanced Neurobiology II - CourseraДокумент5 страницAdvanced Neurobiology II - CourseraSinisa RisticОценок пока нет
- Sleep - Neurobiology, Medicine, and Society - CourseraДокумент6 страницSleep - Neurobiology, Medicine, and Society - CourseraSinisa Ristic0% (1)
- PinterestДокумент4 страницыPinterestSinisa RisticОценок пока нет
- Drugs and The Brain - CourseraДокумент5 страницDrugs and The Brain - CourseraSinisa RisticОценок пока нет
- Advanced Neurobiology II - CourseraДокумент5 страницAdvanced Neurobiology II - CourseraSinisa RisticОценок пока нет
- 10.10-10.30-2019 Seminar On Sports Medicine For Developing CountriesДокумент3 страницы10.10-10.30-2019 Seminar On Sports Medicine For Developing CountriesSinisa RisticОценок пока нет
- 1010-1030-Seminar For Education Managers From BRI CountriesДокумент3 страницы1010-1030-Seminar For Education Managers From BRI CountriesSinisa RisticОценок пока нет
- 2015 Program Kineskog SeminaraДокумент16 страниц2015 Program Kineskog SeminaraSinisa RisticОценок пока нет
- Cirrincione 2019Документ45 страницCirrincione 2019Sinisa RisticОценок пока нет
- (Mebooksfree Net) Dia&ult&phy&equ&hos&mar&thr&3rd PDFДокумент401 страница(Mebooksfree Net) Dia&ult&phy&equ&hos&mar&thr&3rd PDFSinisa RisticОценок пока нет
- 1107-1120 - Seminar On Big Data Application Impact and Telecom Business AnalysisДокумент3 страницы1107-1120 - Seminar On Big Data Application Impact and Telecom Business AnalysisSinisa RisticОценок пока нет
- Section 1Документ34 страницыSection 1Sinisa RisticОценок пока нет
- Clinical and Vaccine Immunology-2007-Taudorf-250.fullДокумент6 страницClinical and Vaccine Immunology-2007-Taudorf-250.fullSinisa RisticОценок пока нет
- Fnmol 11 00216Документ13 страницFnmol 11 00216Sinisa RisticОценок пока нет
- Physrev 00036 2015Документ34 страницыPhysrev 00036 2015Sinisa RisticОценок пока нет
- Nonverbal CommunicationДокумент5 страницNonverbal CommunicationkalimeriОценок пока нет
- Section 1Документ34 страницыSection 1Sinisa RisticОценок пока нет
- Аs 23 10 2019 finalДокумент13 страницАs 23 10 2019 finalSinisa RisticОценок пока нет
- A Meta-AnalysisДокумент9 страницA Meta-AnalysisSinisa RisticОценок пока нет
- ALB2Документ4 страницыALB2Jonalyn SalandoОценок пока нет
- 0 AlignMeДокумент6 страниц0 AlignMeCRISTIAN GABRIEL ZAMBRANO VEGAОценок пока нет
- LPL - Amit Sharma FPSC Loni Road-Gzb Shop No-4, Saharanpur Road, Loni Road, Main Tiraha, Loni, Ghaziabad, Loni S.OДокумент3 страницыLPL - Amit Sharma FPSC Loni Road-Gzb Shop No-4, Saharanpur Road, Loni Road, Main Tiraha, Loni, Ghaziabad, Loni S.OKaran PrabhakarОценок пока нет
- ABCD Vocal Technique Handout PDFДокумент9 страницABCD Vocal Technique Handout PDFRanieque RamosОценок пока нет
- TGD FrameworkДокумент10 страницTGD FrameworkMatthew KleeОценок пока нет
- Jagaroo & Santangelo - Neurophenotypes Advancing Psychiatry and Neuropsychology 2016Документ305 страницJagaroo & Santangelo - Neurophenotypes Advancing Psychiatry and Neuropsychology 2016Jaime Fernández-Aguirrebengoa100% (1)
- Chapter 1Документ4 страницыChapter 1Nikoleta RudnikОценок пока нет
- Tushar FinalДокумент29 страницTushar FinalRaj Prixit RathoreОценок пока нет
- Life Span: HumanДокумент738 страницLife Span: HumanWAJEEHA GOHARОценок пока нет
- Muscle Biopsy A Diagnostic Tool in Muscle DiseasesДокумент9 страницMuscle Biopsy A Diagnostic Tool in Muscle DiseasesRosa AquinoОценок пока нет
- Multiple AlleleДокумент3 страницыMultiple AlleleAdroit SangmaОценок пока нет
- CROSS TREE - Sorte I Selekcije Voćaka I Vinove Loze Iz Severozapadne Srbije I Istočne Hrvatske PDFДокумент153 страницыCROSS TREE - Sorte I Selekcije Voćaka I Vinove Loze Iz Severozapadne Srbije I Istočne Hrvatske PDFrraattaarrrraattaaОценок пока нет
- The 12 Untapped Targets To Ignite New Muscle GrowthДокумент68 страницThe 12 Untapped Targets To Ignite New Muscle Growthace_mimОценок пока нет
- An Examination and Critique of Current Methods To Determine Exercise IntensityДокумент28 страницAn Examination and Critique of Current Methods To Determine Exercise IntensityPabloAñonОценок пока нет
- Masters Portfolio - MSC EmTech (AA, London)Документ56 страницMasters Portfolio - MSC EmTech (AA, London)asad nasirОценок пока нет
- Noncanonical Transnitrosylation Network Contributes To Synapse Loss in Alzheimer's DiseaseДокумент18 страницNoncanonical Transnitrosylation Network Contributes To Synapse Loss in Alzheimer's DiseasedmitworОценок пока нет
- EASL 2021 Version 4 NewДокумент691 страницаEASL 2021 Version 4 NewGupse Köroğlu AdalıОценок пока нет
- Hierarchy ExampleДокумент18 страницHierarchy ExampleSanket SahuОценок пока нет
- NutritionДокумент5 страницNutritionk,srikanthОценок пока нет
- Tomasello, M. (2000) - Culture and Cognitive Development.Документ4 страницыTomasello, M. (2000) - Culture and Cognitive Development.Roberto OcampoОценок пока нет
- Short Note Biology Form 5-Chapter 3 Coordination and ResponseДокумент6 страницShort Note Biology Form 5-Chapter 3 Coordination and Responsesalamah_sabri75% (4)
- Embryo Development (Embryogenesis)Документ31 страницаEmbryo Development (Embryogenesis)SHEMAR STEWARTОценок пока нет
- Grade 5, Science CurriculumДокумент3 страницыGrade 5, Science CurriculumJoel C. Yuvienco67% (3)
- Kaid Seloua-PdbДокумент9 страницKaid Seloua-Pdbseloua kaidОценок пока нет
- Characterization of Tannia PlantДокумент104 страницыCharacterization of Tannia PlantSolomon FantawОценок пока нет
- Cell and Tissue Culture: Model Cells in Medicine - Stem Cells - Tissue EngineeringДокумент44 страницыCell and Tissue Culture: Model Cells in Medicine - Stem Cells - Tissue EngineeringCrasilia Yanti PadangОценок пока нет
- Can The Unconscious Mind Be Persuaded? An Overview With Marketing ImplicationsДокумент9 страницCan The Unconscious Mind Be Persuaded? An Overview With Marketing ImplicationsAhmed El-ShafeiОценок пока нет
- Social Institutions - Kinship, Family&Marriage PDFДокумент47 страницSocial Institutions - Kinship, Family&Marriage PDFAbhijith KОценок пока нет
- The Tai Chi Code - Martial Arts Ebook by Chris DavisДокумент190 страницThe Tai Chi Code - Martial Arts Ebook by Chris DavisTomas De Wael100% (2)
- LeaP Science 10 Q4 W3 4Документ5 страницLeaP Science 10 Q4 W3 4D Garcia100% (2)
- Gut: the new and revised Sunday Times bestsellerОт EverandGut: the new and revised Sunday Times bestsellerРейтинг: 4 из 5 звезд4/5 (392)
- Tales from Both Sides of the Brain: A Life in NeuroscienceОт EverandTales from Both Sides of the Brain: A Life in NeuroscienceРейтинг: 3 из 5 звезд3/5 (18)
- Why We Die: The New Science of Aging and the Quest for ImmortalityОт EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityРейтинг: 4 из 5 звезд4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisОт EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisРейтинг: 3.5 из 5 звезд3.5/5 (2)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)От EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Рейтинг: 4 из 5 звезд4/5 (378)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperОт EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperРейтинг: 4.5 из 5 звезд4.5/5 (15)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceОт EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceРейтинг: 4.5 из 5 звезд4.5/5 (516)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouОт EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouРейтинг: 4.5 из 5 звезд4.5/5 (62)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsОт EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsРейтинг: 4 из 5 звезд4/5 (5)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessОт Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessРейтинг: 4 из 5 звезд4/5 (33)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionОт EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionРейтинг: 4 из 5 звезд4/5 (811)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedОт EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedРейтинг: 4 из 5 звезд4/5 (11)
- Human: The Science Behind What Makes Your Brain UniqueОт EverandHuman: The Science Behind What Makes Your Brain UniqueРейтинг: 3.5 из 5 звезд3.5/5 (38)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesОт EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesРейтинг: 4.5 из 5 звезд4.5/5 (397)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorОт EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorОценок пока нет
- Who's in Charge?: Free Will and the Science of the BrainОт EverandWho's in Charge?: Free Will and the Science of the BrainРейтинг: 4 из 5 звезд4/5 (65)
- Good Without God: What a Billion Nonreligious People Do BelieveОт EverandGood Without God: What a Billion Nonreligious People Do BelieveРейтинг: 4 из 5 звезд4/5 (66)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldОт EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldРейтинг: 4.5 из 5 звезд4.5/5 (18)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondОт EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondРейтинг: 4 из 5 звезд4/5 (4)
- Inside of a Dog: What Dogs See, Smell, and KnowОт EverandInside of a Dog: What Dogs See, Smell, and KnowРейтинг: 4 из 5 звезд4/5 (390)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemОт EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemРейтинг: 4.5 из 5 звезд4.5/5 (115)
- The Invention of Tomorrow: A Natural History of ForesightОт EverandThe Invention of Tomorrow: A Natural History of ForesightРейтинг: 4.5 из 5 звезд4.5/5 (5)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomОт EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomРейтинг: 4 из 5 звезд4/5 (215)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineОт EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineРейтинг: 4 из 5 звезд4/5 (17)