Вам также может понравиться
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- Oral Lec 8Документ9 страницOral Lec 8Adam AliraqiОценок пока нет
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Laboratory Exercise 1Документ2 страницыLaboratory Exercise 1Markie Dela CruzОценок пока нет
- AP1 Lab4 Intro To Skeleton FA2021Документ15 страницAP1 Lab4 Intro To Skeleton FA2021Kim VenturaОценок пока нет
- Management of Severe Hyperkalemia PDFДокумент6 страницManagement of Severe Hyperkalemia PDFCarlos Navarro YslaОценок пока нет
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- 4.prenatal and Postnatal Development of MandibleДокумент77 страниц4.prenatal and Postnatal Development of Mandibleakshay100% (2)
- Enteric Nervous SystemДокумент12 страницEnteric Nervous SystemedgarОценок пока нет
- Physical Assessment Vital SignsДокумент17 страницPhysical Assessment Vital SignsArvi GayeОценок пока нет
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- Materi HE GinjalДокумент61 страницаMateri HE GinjalYafi Dyah C.IОценок пока нет
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Histology - Respiratory SystemДокумент14 страницHistology - Respiratory SystemAbiola NerdОценок пока нет
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Dental MCQДокумент22 страницыDental MCQPinpointq0% (1)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- 47pages METHAMPHETAMINE TOXICITY AND MESSENGERS OF DEATHДокумент47 страниц47pages METHAMPHETAMINE TOXICITY AND MESSENGERS OF DEATHDenise MathreОценок пока нет
- ABG ElectrolytesДокумент48 страницABG ElectrolytesDRwaqas Gulzar100% (1)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- April PhysiologyДокумент2 страницыApril Physiologyapi-284800663Оценок пока нет
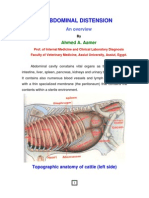
- Abdominal DistensionДокумент10 страницAbdominal DistensionMorad ImadОценок пока нет
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- Yuvdz3wu 13jДокумент2 страницыYuvdz3wu 13jNael LacerdaОценок пока нет
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- Assessment Nursing Diagnosis Analysis Goals and Objectives Nursing Interventions Rationale Evaluation EffectivenessДокумент3 страницыAssessment Nursing Diagnosis Analysis Goals and Objectives Nursing Interventions Rationale Evaluation EffectivenessYnah Sayoc75% (4)
- Biomechanics in Applications PDFДокумент424 страницыBiomechanics in Applications PDFRAUL EDUARDO GUTIERREZ COITIÑO100% (1)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- Toxic A History of Nerve Agents From Nazi Germany To Putins Russia 1St Edition Edition Dan Kaszeta Download 2024 Full ChapterДокумент23 страницыToxic A History of Nerve Agents From Nazi Germany To Putins Russia 1St Edition Edition Dan Kaszeta Download 2024 Full Chaptermari.artrip178100% (9)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- Hepatorenal Syndrome: Pathophysiologic Basis of Therapy and Current ManagementДокумент93 страницыHepatorenal Syndrome: Pathophysiologic Basis of Therapy and Current ManagementJOHN ALFREY PUEBLOОценок пока нет
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- COVID ProlongadoДокумент22 страницыCOVID ProlongadoSMIBA MedicinaОценок пока нет
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- HypothermiaДокумент4 страницыHypothermiaGerald YasonОценок пока нет
- Digestion Test BiologyДокумент3 страницыDigestion Test BiologyKatrīna SimanovskaОценок пока нет
- BISC403 Sample Exam 4 W - Answers 20SpДокумент5 страницBISC403 Sample Exam 4 W - Answers 20SpGrace MillsОценок пока нет
- Anatomy & Physiology of Olfactory System.Документ27 страницAnatomy & Physiology of Olfactory System.Prasanna DattaОценок пока нет
- Good BooksДокумент16 страницGood BooksFbihansip bancel100% (1)
- IIIДокумент13 страницIIIapi-3828211Оценок пока нет
- Feliciano, Troy Ivan Silva, Queenie Rose V. BSN 3-C: Rationale: To Track The Changes of Client's ConditionДокумент4 страницыFeliciano, Troy Ivan Silva, Queenie Rose V. BSN 3-C: Rationale: To Track The Changes of Client's ConditionQueenie SilvaОценок пока нет
- The MoneraДокумент19 страницThe MoneraNaufal Taufiq AmmarОценок пока нет
- Talley & O'Connor Quiz SampleДокумент5 страницTalley & O'Connor Quiz SamplefilchibuffОценок пока нет
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Case Report Myoma Uterine: Supervised By: Dr. Hesty Duhita Permata, SP - OGДокумент12 страницCase Report Myoma Uterine: Supervised By: Dr. Hesty Duhita Permata, SP - OGNiki Rizqi rachmawatiОценок пока нет