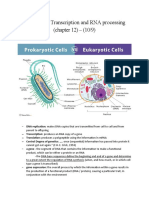
Вам также может понравиться
- VCE Biology ExamДокумент29 страницVCE Biology ExamVy PhanОценок пока нет
- Gene RegulationДокумент30 страницGene Regulationprehealthhelp67% (3)
- Reviewer in Science 10Документ2 страницыReviewer in Science 10fan girlОценок пока нет
- Central DogmaДокумент50 страницCentral DogmaIvilyn ManguladОценок пока нет
- Devlin Biochemistry With Clinical CorrelationsДокумент1 241 страницаDevlin Biochemistry With Clinical CorrelationsJorge100% (5)
- MRCP Notes 2006Документ117 страницMRCP Notes 2006Walaa Ismail HidirbiОценок пока нет
- Artikel 4Документ15 страницArtikel 4Angelina KobanОценок пока нет
- BIO 310 Midterm 2 + Final Exam PackageДокумент44 страницыBIO 310 Midterm 2 + Final Exam PackageNerdy Notes Inc.Оценок пока нет
- Regulatory RNAs in BacteriaДокумент14 страницRegulatory RNAs in BacteriaDaniel Alberto Bothia MartinezОценок пока нет
- Riboswitches Control Fundamental Biochemical Pathways in Bacillus Subtilis and Other BacteriaДокумент10 страницRiboswitches Control Fundamental Biochemical Pathways in Bacillus Subtilis and Other BacteriarednriОценок пока нет
- Proteinas AbcteriasДокумент15 страницProteinas AbcteriasCindy GarciaОценок пока нет
- RNA Interference Class NotesДокумент4 страницыRNA Interference Class NotesmanaleepandyaОценок пока нет
- D Mallika ComputerscienceДокумент62 страницыD Mallika ComputerscienceDhilsanth SLОценок пока нет
- Castañeda 2016Документ16 страницCastañeda 2016papahojaloveОценок пока нет
- Albertomaria Moro Thesis Final VersionДокумент97 страницAlbertomaria Moro Thesis Final VersionGergő BárdosОценок пока нет
- Regulation of Gene Expression PDFДокумент4 страницыRegulation of Gene Expression PDFPranav Kumar MishraОценок пока нет
- Epigenética y ARNДокумент9 страницEpigenética y ARNFrank BustosОценок пока нет
- Traduccuón en ProcariotasДокумент21 страницаTraduccuón en Procariotasmaria fernanda jarquin marquezОценок пока нет
- Emerging Roles of Rna-Binding Proteins in Plant Development: SciencedirectДокумент7 страницEmerging Roles of Rna-Binding Proteins in Plant Development: SciencedirectPavithra MohanОценок пока нет
- 1 s2.0 S0968000415001644 MainДокумент11 страниц1 s2.0 S0968000415001644 MainTheresa LambayonОценок пока нет
- Epigenetics Role in Alternative RNA SplicingДокумент11 страницEpigenetics Role in Alternative RNA SplicingThiagoОценок пока нет
- Modifications Controlling RNAДокумент5 страницModifications Controlling RNADiego RodriguezОценок пока нет
- RNA SplicingДокумент14 страницRNA SplicingnurlianaОценок пока нет
- E2017005 PDFДокумент6 страницE2017005 PDFComputational and Structural Biotechnology JournalОценок пока нет
- 9 J Pathology 2010 AДокумент14 страниц9 J Pathology 2010 Amilenerato2240Оценок пока нет
- Ekspresi Gen PDFДокумент12 страницEkspresi Gen PDFRose DesyОценок пока нет
- Fis Upstream BindingДокумент15 страницFis Upstream BindingDiegoОценок пока нет
- Widespread Generation of Alternative UTRs Contributes To Sex-Specific RNA Binding by UNRДокумент12 страницWidespread Generation of Alternative UTRs Contributes To Sex-Specific RNA Binding by UNRWasim AОценок пока нет
- REVISED - RNAi Circuit ManuscriptДокумент20 страницREVISED - RNAi Circuit ManuscriptOscar PellyОценок пока нет
- Seminario 3.2 - Micro RNA - Biogenesis, Mechanism of Action and Applications-A ReviewДокумент27 страницSeminario 3.2 - Micro RNA - Biogenesis, Mechanism of Action and Applications-A ReviewAngellique NiyiragiraОценок пока нет
- Gene Regulation of Mammalian Long Non Coding RNA: Heeyoun BunchДокумент15 страницGene Regulation of Mammalian Long Non Coding RNA: Heeyoun Bunchmehdi_mohebi2009Оценок пока нет
- Transcription Factor Dynamics Mini ReviewДокумент8 страницTranscription Factor Dynamics Mini ReviewwebhappyОценок пока нет
- Ma1 PDFДокумент21 страницаMa1 PDFJoyceline DavidОценок пока нет
- Cells 10 01831 v2Документ24 страницыCells 10 01831 v2msiddique13216Оценок пока нет
- Transcription FactorsДокумент25 страницTranscription FactorsPriya.RОценок пока нет
- Genome-Wide Analyses Across Viridiplantae Reveal TДокумент12 страницGenome-Wide Analyses Across Viridiplantae Reveal TVictor CostaОценок пока нет
- Marchese Genome Biol 2017 - Rev lncRNAДокумент13 страницMarchese Genome Biol 2017 - Rev lncRNABrahim León DailalОценок пока нет
- Unit 3Документ33 страницыUnit 3Dsce BtОценок пока нет
- Identification and Characterization of Long Non-Coding RNA and Their Response Against Citrus Bark Cracking Viroid Infection in Humulus LupulusДокумент15 страницIdentification and Characterization of Long Non-Coding RNA and Their Response Against Citrus Bark Cracking Viroid Infection in Humulus LupulusLukas MichelОценок пока нет
- Regulation of Gene Expression - OurvleДокумент17 страницRegulation of Gene Expression - Ourvledpkf6dj725Оценок пока нет
- Rnase Fused ProteinДокумент14 страницRnase Fused ProteinYanjiaoОценок пока нет
- A Symphony of Transcription Factors For Gene ControlДокумент19 страницA Symphony of Transcription Factors For Gene ControlEdgardo Becerra BecerraОценок пока нет
- McCown Et Al. MiMB 2012Документ17 страницMcCown Et Al. MiMB 2012Phil McCownОценок пока нет
- 1 s2.0 S1874939919302160 MainДокумент11 страниц1 s2.0 S1874939919302160 MainxОценок пока нет
- Mechanisms and Functions of Long Non-Coding RNAs AДокумент29 страницMechanisms and Functions of Long Non-Coding RNAs AClaudio Andres Lamilla MardonesОценок пока нет
- Chapter 11 - Gene ExpressionДокумент2 страницыChapter 11 - Gene Expressionapi-272677238Оценок пока нет
- Ghaz-Jahanian2013 Article InfluenceOfSmallRNAsOnBiofilmFДокумент10 страницGhaz-Jahanian2013 Article InfluenceOfSmallRNAsOnBiofilmFpapahojaloveОценок пока нет
- Chen (2020)Документ15 страницChen (2020)IVAN LUIS FERNANDEZ DE LA CRUZОценок пока нет
- Mechanism & FunctionsДокумент7 страницMechanism & FunctionsShaher Bano MirzaОценок пока нет
- Lecture 2 - Transcription and RNA ProcessingДокумент22 страницыLecture 2 - Transcription and RNA ProcessingErika KulićОценок пока нет
- RNAi MphilДокумент28 страницRNAi Mphilrameeshasaleem01Оценок пока нет
- RNAi in Developmental BiologyДокумент7 страницRNAi in Developmental BiologyAaryan GuptaОценок пока нет
- Anatomy of A GeneДокумент33 страницыAnatomy of A GenemskikiОценок пока нет
- Understanding Mechanisms of Gene Expression: E. LosДокумент2 страницыUnderstanding Mechanisms of Gene Expression: E. LosRuchi SharmaОценок пока нет
- TECHNIQUES IN MOLECULAR BIOLOGY – PROMOTERS AND PLASMIDSДокумент4 страницыTECHNIQUES IN MOLECULAR BIOLOGY – PROMOTERS AND PLASMIDSAndika MardiantoОценок пока нет
- Microbiology 2421 Lecture Notes Microbial GeneticsДокумент19 страницMicrobiology 2421 Lecture Notes Microbial GeneticsFaye Coleen GozaОценок пока нет
- Rna InterferenceДокумент10 страницRna InterferencewaktoleОценок пока нет
- 2010 Comparative Genomics Reveals 104 Candidate Structured RNAs From Bacteria, Archaea, and Their MetagenomesДокумент17 страниц2010 Comparative Genomics Reveals 104 Candidate Structured RNAs From Bacteria, Archaea, and Their Metagenomesplanhigion06Оценок пока нет
- 2020 in Vitro Ribosome Synthesis and Evolution Through Ribosome DisplayДокумент10 страниц2020 in Vitro Ribosome Synthesis and Evolution Through Ribosome DisplayLai weng yu Lai weng yuОценок пока нет
- Molecular Biology NotesДокумент8 страницMolecular Biology Notesbyunus88Оценок пока нет
- 1 s2.0 S1931312819305281 MainДокумент25 страниц1 s2.0 S1931312819305281 MainJorge Lima PérezОценок пока нет
- Assignment-04 22 23Документ1 страницаAssignment-04 22 23Gi GiОценок пока нет
- Analytical BiochemistryДокумент7 страницAnalytical BiochemistryCelssy NataliОценок пока нет
- Controlofgeneexpression 180801122812Документ46 страницControlofgeneexpression 180801122812owadokunolamipoОценок пока нет
- The RNA-induced Silencing Complex - A Versatile Gene-Silencing MachineДокумент5 страницThe RNA-induced Silencing Complex - A Versatile Gene-Silencing MachineWillОценок пока нет
- Malassezia Pachydermatis: Carriage in Dog OwnersДокумент6 страницMalassezia Pachydermatis: Carriage in Dog OwnershansmeetОценок пока нет
- Immunotherapy in Invasive Fungal Diseases 2015Документ10 страницImmunotherapy in Invasive Fungal Diseases 2015hansmeetОценок пока нет
- Neurocysticercosis: A Review: January 2007Документ6 страницNeurocysticercosis: A Review: January 2007hansmeetОценок пока нет
- 6.3 Medical Mycology and Fungal ImmunologyДокумент10 страниц6.3 Medical Mycology and Fungal ImmunologyhansmeetОценок пока нет
- Neurocysticercosis: A Disease of Neglect: Abhishek Mewara, Kapil Goyal, Rakesh SehgalДокумент9 страницNeurocysticercosis: A Disease of Neglect: Abhishek Mewara, Kapil Goyal, Rakesh SehgalhansmeetОценок пока нет
- 5.4 Novel Approaches Diagnostic Mycology 2019Документ8 страниц5.4 Novel Approaches Diagnostic Mycology 2019hansmeetОценок пока нет
- Nosocomial Infections and Zoonotic Diseases in Veterinary HospitalsДокумент4 страницыNosocomial Infections and Zoonotic Diseases in Veterinary HospitalshansmeetОценок пока нет
- Priorities For Research and Control of Cestode Zoonoses in AsiaДокумент11 страницPriorities For Research and Control of Cestode Zoonoses in AsiahansmeetОценок пока нет
- Nonculture Methods For Diagnosis 2002Документ20 страницNonculture Methods For Diagnosis 2002hansmeetОценок пока нет
- Chapitre Surveillance GeneralДокумент10 страницChapitre Surveillance GeneralhansmeetОценок пока нет
- 5.3 Curr Challenges Diagnosis of Fungal Infs 2017Документ13 страниц5.3 Curr Challenges Diagnosis of Fungal Infs 2017hansmeetОценок пока нет
- Chou Dhuri 2012Документ10 страницChou Dhuri 2012yakoub bensaadaОценок пока нет
- Nosocomial Infections and Zoonotic Diseases in Veterinary HospitalsДокумент4 страницыNosocomial Infections and Zoonotic Diseases in Veterinary HospitalshansmeetОценок пока нет
- Kuntsi Vaattovaara2002 PDFДокумент6 страницKuntsi Vaattovaara2002 PDFhansmeetОценок пока нет
- Laboratory Diagnosis of The Causative Dermatophytes of Tinea CapitisДокумент15 страницLaboratory Diagnosis of The Causative Dermatophytes of Tinea CapitisMohamed KhalidОценок пока нет
- Risks of Nosocomial Zoonotic TransmissionДокумент11 страницRisks of Nosocomial Zoonotic TransmissionhansmeetОценок пока нет
- Recent Advances in The Diagnosis of DermatophytosisДокумент11 страницRecent Advances in The Diagnosis of DermatophytosishansmeetОценок пока нет
- Bacterial Protein Toxins That Modify Host Regulatory GTPases PDFДокумент12 страницBacterial Protein Toxins That Modify Host Regulatory GTPases PDFKraken UrОценок пока нет
- Shelter Hospital Mode: How Do We Prevent COVID-19 Hospital-Acquired Infection?Документ2 страницыShelter Hospital Mode: How Do We Prevent COVID-19 Hospital-Acquired Infection?hansmeetОценок пока нет
- Challenges in The Development of Effective Peptide Vaccines For CancerДокумент11 страницChallenges in The Development of Effective Peptide Vaccines For CancerhansmeetОценок пока нет
- Design of A Recombinant Hepatitis B Vaccine Based On Stably Binding Hlai Peptides 2167 7956.1000120Документ4 страницыDesign of A Recombinant Hepatitis B Vaccine Based On Stably Binding Hlai Peptides 2167 7956.1000120hansmeetОценок пока нет
- Old and New Vaccine Approaches: Ruth Arnon, Tamar Ben-YedidiaДокумент10 страницOld and New Vaccine Approaches: Ruth Arnon, Tamar Ben-YedidiahansmeetОценок пока нет
- Rotary Motor of Bacterial PDFДокумент38 страницRotary Motor of Bacterial PDFhansmeetОценок пока нет
- Approaches To Improve Chemically Defined Synthetic Peptide VaccinesДокумент8 страницApproaches To Improve Chemically Defined Synthetic Peptide VaccineshansmeetОценок пока нет
- 1 Gene Regulation by Riboswitches PDFДокумент13 страниц1 Gene Regulation by Riboswitches PDFhansmeetОценок пока нет
- What Is A Vaccine?: Technical Notes (/protein-Technical-Notes - HTML?SRC Leftbar)Документ4 страницыWhat Is A Vaccine?: Technical Notes (/protein-Technical-Notes - HTML?SRC Leftbar)hansmeetОценок пока нет
- 1 Bacterial Toxins PDFДокумент30 страниц1 Bacterial Toxins PDFhansmeetОценок пока нет
- 1 Bacterial InvasionДокумент13 страниц1 Bacterial InvasionhansmeetОценок пока нет
- 1 Bacterial InvasionДокумент13 страниц1 Bacterial InvasionhansmeetОценок пока нет
- LECTURE 9-Transgenic Technology in Marine Organisms PDFДокумент26 страницLECTURE 9-Transgenic Technology in Marine Organisms PDFIntan Lestari DewiОценок пока нет
- BIO Genetics Eukaryote TranscriptionДокумент23 страницыBIO Genetics Eukaryote TranscriptionAnonymous SVy8sOsvJDОценок пока нет
- B. Exon: C. Transcription, TranslationДокумент5 страницB. Exon: C. Transcription, TranslationRuhna BaigОценок пока нет
- Xii-Biology Cbse Revision 2Документ10 страницXii-Biology Cbse Revision 2pavithra KumarОценок пока нет
- Gen Bio Mock QuizДокумент6 страницGen Bio Mock QuizJarvis RazonОценок пока нет
- NEET Biology Test Questions on Molecular BiologyДокумент4 страницыNEET Biology Test Questions on Molecular BiologySadananda KumbhakarОценок пока нет
- AMC Test Sample PaperДокумент36 страницAMC Test Sample PaperArsalan HussainОценок пока нет
- Human Biology: Pearson Edexcel International GCSE (9-1)Документ24 страницыHuman Biology: Pearson Edexcel International GCSE (9-1)Nang Seng Tawng HparawОценок пока нет
- Transgenic Fly FinalДокумент7 страницTransgenic Fly FinalJOHN THRONN DE JESUS100% (1)
- Transcription ProcessДокумент85 страницTranscription Processshruti shahОценок пока нет
- Test Bank For Biology 12th by RavenДокумент52 страницыTest Bank For Biology 12th by RavenAmberClineagwbd100% (38)
- Q3-W4B-Protein Synthesis (By Group)Документ5 страницQ3-W4B-Protein Synthesis (By Group)Catherine CantadaОценок пока нет
- Dwnload Full Beckers World of The Cell 9th Edition Hardin Solutions Manual PDFДокумент35 страницDwnload Full Beckers World of The Cell 9th Edition Hardin Solutions Manual PDFgebbielean1237100% (12)
- BCH 305 (Chemistry & Metabolism of Nucleic Acids)Документ30 страницBCH 305 (Chemistry & Metabolism of Nucleic Acids)ihebunnaogochukwu24Оценок пока нет
- Mitochondrial & Chloroplast Genomes Circular Under 50kbДокумент5 страницMitochondrial & Chloroplast Genomes Circular Under 50kbbmhshОценок пока нет
- Cambridge IGCSE™: Biology 0610/42Документ12 страницCambridge IGCSE™: Biology 0610/42Mahir VasoyaОценок пока нет
- Protein Synthesis (Translation) : IlosДокумент14 страницProtein Synthesis (Translation) : Ilosghost beastОценок пока нет
- Molecular Basis Off InheritanceДокумент5 страницMolecular Basis Off InheritanceShivam Kumar PathakОценок пока нет
- محاضرة 2Документ41 страницаمحاضرة 2AbdulОценок пока нет
- Lecture Unit 3 Science Grade 10Документ6 страницLecture Unit 3 Science Grade 10Krizt Theodore Colico100% (7)
- Study Guide Dna Rna No VirusДокумент4 страницыStudy Guide Dna Rna No VirusLara AlbeeshyОценок пока нет
- CH 12Документ40 страницCH 12abdurОценок пока нет
- Crop LifeДокумент7 страницCrop Life유가연[학생](생명과학대학 유전생명공학과)Оценок пока нет
- (Advances in Parasitology 76) Louis M. Weiss and Herbert B. Tanowitz (Eds.) - Chagas Disease, Part B-Academic Press, Elsevier (2011)Документ283 страницы(Advances in Parasitology 76) Louis M. Weiss and Herbert B. Tanowitz (Eds.) - Chagas Disease, Part B-Academic Press, Elsevier (2011)Stoian GoranovОценок пока нет