Вам также может понравиться
- Teoría polivagal: Descubrir el poder curativo del nervio vago para superar la ansiedad, el trauma, la depresión y el estrés emocionalОт EverandTeoría polivagal: Descubrir el poder curativo del nervio vago para superar la ansiedad, el trauma, la depresión y el estrés emocionalРейтинг: 4 из 5 звезд4/5 (6)
- Las Fracturas de Columna VertebralДокумент5 страницLas Fracturas de Columna VertebralJuan Carlos Claros OliveraОценок пока нет
- Psicooncología: Ayuda Psicológica Para Pacientes Con CáncerОт EverandPsicooncología: Ayuda Psicológica Para Pacientes Con CáncerРейтинг: 4 из 5 звезд4/5 (1)
- Consejos para Después Del Alisado de KeratinaДокумент2 страницыConsejos para Después Del Alisado de KeratinaramseyptvaoxfixjОценок пока нет
- EquinoculturaДокумент16 страницEquinoculturafiocelabeОценок пока нет
- 2012 - Caso Clínico Evaluación e Intervención en Un Caso de Ansiedad Por Separación de Una Niña Adoptada - Fdez-Zúñiga Marcos PDFДокумент11 страниц2012 - Caso Clínico Evaluación e Intervención en Un Caso de Ansiedad Por Separación de Una Niña Adoptada - Fdez-Zúñiga Marcos PDFskinner2011Оценок пока нет
- La pizarra de Babel: Puentes entre neurociencia, psicología y educaciónОт EverandLa pizarra de Babel: Puentes entre neurociencia, psicología y educaciónРейтинг: 5 из 5 звезд5/5 (1)
- Clasificacion para Terceros Molares2Документ8 страницClasificacion para Terceros Molares2Alejandro MorenoОценок пока нет
- Cómo se comunican las neuronas: El milagro de la transmisión sinápticaОт EverandCómo se comunican las neuronas: El milagro de la transmisión sinápticaОценок пока нет
- 1 Caso Sindrome PrefrontalДокумент5 страниц1 Caso Sindrome PrefrontalBroken fallОценок пока нет
- Funciones Cerebrales Superiores SemiologДокумент8 страницFunciones Cerebrales Superiores SemiologZuirz Mijoha100% (1)
- Regulacion EmocionalДокумент8 страницRegulacion EmocionalAzul MerloОценок пока нет
- La Organización Funcional Del CerebroДокумент13 страницLa Organización Funcional Del CerebroJosé Rodrigo Torrico CrespoОценок пока нет
- Síndromes prefrontales: funciones y lesionesДокумент9 страницSíndromes prefrontales: funciones y lesionesAzul MerloОценок пока нет
- Plasticidad CerebralДокумент19 страницPlasticidad CerebralIbarra MarthaОценок пока нет
- Caso Clínico N - 3 NeuroДокумент4 страницыCaso Clínico N - 3 NeuroAngelo Baeza SprohnleОценок пока нет
- Movimientos MorfogeneticosДокумент2 страницыMovimientos MorfogeneticosAlberto Lira100% (6)
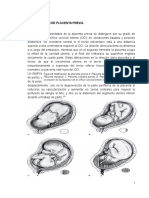
- Clasificación de Placenta PreviaДокумент2 страницыClasificación de Placenta PreviaKelys R. Padilla100% (1)
- Proyecto de Investigacion Sobre El SuicidioДокумент44 страницыProyecto de Investigacion Sobre El Suicidiolucia oliveiraОценок пока нет
- Los Lobulos Frontales y Las Funciones Ej PDFДокумент93 страницыLos Lobulos Frontales y Las Funciones Ej PDFPío Augusto Alvarez ValenciaОценок пока нет
- SX Frontal Medial SuperiorДокумент9 страницSX Frontal Medial SuperiorLucio Alexánder Acero RodríguezОценок пока нет
- Base de La Autorrepresentacion y Sus Alteraciones Patológicas PDFДокумент3 страницыBase de La Autorrepresentacion y Sus Alteraciones Patológicas PDFPacoОценок пока нет
- Bases NuropsicologicasДокумент3 страницыBases NuropsicologicasPacoОценок пока нет
- La Cognición Social y La Corteza CerebralДокумент6 страницLa Cognición Social y La Corteza CerebralMariel NaumazevОценок пока нет
- Diálogo Entre Corteza y Subcorteza CerebralДокумент16 страницDiálogo Entre Corteza y Subcorteza CerebralRacingjuliusОценок пока нет
- SX Prefrontales PDFДокумент5 страницSX Prefrontales PDFThalii MorfinОценок пока нет
- 47390110Документ5 страниц47390110Alex Pachas GradosОценок пока нет
- Cognición social y corteza cerebralДокумент7 страницCognición social y corteza cerebralAlan AldreteОценок пока нет
- Funciones de La Corteza Prefrontal Ventromedial en La Toma de Decisiones EmocionalesДокумент30 страницFunciones de La Corteza Prefrontal Ventromedial en La Toma de Decisiones EmocionalesAna Maria RomeroОценок пока нет
- Rains - 2004 - Principios - de - Neuro Cap9Документ27 страницRains - 2004 - Principios - de - Neuro Cap9Ayelen SosaОценок пока нет
- Neuropsicologia Asperger y Sindrome DisejecutivoДокумент13 страницNeuropsicologia Asperger y Sindrome DisejecutivoMariana OliverОценок пока нет
- Ensayo PDFДокумент5 страницEnsayo PDFEmely ObrienОценок пока нет
- Ensayo Sobre Los Contenidos de La ConcienciaДокумент5 страницEnsayo Sobre Los Contenidos de La ConcienciaEmely ObrienОценок пока нет
- ResumosДокумент4 страницыResumosMicaela TavaresОценок пока нет
- Caso ClínicoДокумент12 страницCaso ClínicoRuth Cobo RosalesОценок пока нет
- Repaso 56Документ63 страницыRepaso 56Arvid KnutОценок пока нет
- Neuroanatomia Del S.H.E y Tipos de S.H.EДокумент19 страницNeuroanatomia Del S.H.E y Tipos de S.H.EKAREN LICETH RIVERA HERNANDEZОценок пока нет
- Funciones cognitivas del lóbulo frontalДокумент5 страницFunciones cognitivas del lóbulo frontalVale DoteОценок пока нет
- Caso Sindrome PreforntalДокумент4 страницыCaso Sindrome PreforntalKendy GonzalesОценок пока нет
- Unidad 1 - Ciclo de La Tarea 1-Estructura Del Trabajo A EntregarДокумент9 страницUnidad 1 - Ciclo de La Tarea 1-Estructura Del Trabajo A Entregarangela cristinaОценок пока нет
- Cerebro Humano EnsayoДокумент4 страницыCerebro Humano EnsayoEsmeraldaGarciaОценок пока нет
- Ensayo Unidad 2Документ3 страницыEnsayo Unidad 2Radelsa Mendoza DominguezОценок пока нет
- Cuerpo Calloso PDFДокумент7 страницCuerpo Calloso PDFKarenОценок пока нет
- Tarea 3 y 4 NeuroДокумент9 страницTarea 3 y 4 NeuroNicole JimenezОценок пока нет
- NT 2021 03Документ5 страницNT 2021 03ROCIO ANTONELA CALCINA SUCAОценок пока нет
- La Cognicion Social y La Corteza CerebralДокумент7 страницLa Cognicion Social y La Corteza CerebralMariana CandiaОценок пока нет
- Esructuras Cerebrales en La Percepción PDFДокумент4 страницыEsructuras Cerebrales en La Percepción PDFJeimmy Garcia MarinОценок пока нет
- Monografia Neurobiologia Maria - Valdueza.tauroniДокумент5 страницMonografia Neurobiologia Maria - Valdueza.tauroniSarahi Bautista SОценок пока нет
- Neuroplasticidad Cerebral en Áreas Occipitales en Adolescentes CiegosДокумент5 страницNeuroplasticidad Cerebral en Áreas Occipitales en Adolescentes CiegosRodrigo Aguirre Báez100% (1)
- Trabajo de Investigacion Plasticidad CerebralДокумент16 страницTrabajo de Investigacion Plasticidad CerebralFrancisco César García MuñozОценок пока нет
- Psiquiatria IntroduccionДокумент12 страницPsiquiatria IntroduccionNayeli LeitrochОценок пока нет
- 3 Plasticidad CerebralДокумент6 страниц3 Plasticidad CerebralLaura LopezОценок пока нет
- Power de Miedo, Defensa y AgresionДокумент10 страницPower de Miedo, Defensa y AgresionMarguerite DurasОценок пока нет
- Biotek DGДокумент12 страницBiotek DGedith dayana gonzalez varelaОценок пока нет
- Tarea 4-Estimulación Cognitiva y PsicosocialДокумент14 страницTarea 4-Estimulación Cognitiva y PsicosocialYekiro DumeОценок пока нет
- Psicoan Lisis y Neurociencias H. BleichmarДокумент7 страницPsicoan Lisis y Neurociencias H. BleichmarIsaac RamírezОценок пока нет
- Caso Clínico 3.0Документ4 страницыCaso Clínico 3.0VentasTemucoPitrufquénОценок пока нет
- Agabriela - Arriaga - Unidad 1 - Actividad 3 - Sistemas CerebralesДокумент4 страницыAgabriela - Arriaga - Unidad 1 - Actividad 3 - Sistemas Cerebralesdaniel chaparroОценок пока нет
- La atención cerebralДокумент9 страницLa atención cerebralAndrea Gallo de la PazОценок пока нет
- Sistema LimbicoДокумент8 страницSistema LimbicoWillians Ramírez leyvaОценок пока нет
- Ao 11-1Документ17 страницAo 11-1Una patata Llamada AntonellaОценок пока нет
- El rápido viaje del miedo en la amígdala cerebralДокумент3 страницыEl rápido viaje del miedo en la amígdala cerebralMaria P. LópezОценок пока нет
- Evaluacion Neuropsic de Meningioma FrotnalДокумент10 страницEvaluacion Neuropsic de Meningioma FrotnalMonaОценок пока нет
- Informe 2 Rogel Nuñez Educacion FisicaДокумент5 страницInforme 2 Rogel Nuñez Educacion FisicaTecno SanОценок пока нет
- Politica Tratamiento de La InformacionДокумент7 страницPolitica Tratamiento de La InformacionMaritza mojicaОценок пока нет
- Protocolo Colocación y Retiro Del EPP Mayo 2020Документ7 страницProtocolo Colocación y Retiro Del EPP Mayo 2020Maritza mojicaОценок пока нет
- OGPSICOLOGIAДокумент5 страницOGPSICOLOGIAMaritza mojicaОценок пока нет
- Formato de Articulo TeincoДокумент20 страницFormato de Articulo TeincoMaritza mojicaОценок пока нет
- Suicidio JovenesДокумент14 страницSuicidio JovenesAnonymous gIfXPkiL9SОценок пока нет
- Segunda Entrega Gestion Social de ProyectosДокумент29 страницSegunda Entrega Gestion Social de ProyectosMaritza mojicaОценок пока нет
- Entrega Uno MetodosДокумент13 страницEntrega Uno MetodosMaritza yadira Mojica avilaОценок пока нет
- Qué Es y Cómo Se Realiza Un EnsayoДокумент4 страницыQué Es y Cómo Se Realiza Un EnsayoJosé Antonio EstrellaОценок пока нет
- Proyecto Psicologia Clinica Primera EntregaДокумент16 страницProyecto Psicologia Clinica Primera EntregaMaritza mojicaОценок пока нет
- Entrega Final Metodos CuantitativosДокумент25 страницEntrega Final Metodos CuantitativosMaritza mojicaОценок пока нет
- Compras y AprovisionamientoДокумент13 страницCompras y AprovisionamientoMaritza yadira Mojica avilaОценок пока нет
- Satisfacción salarial clave permanencia trabajadoresДокумент2 страницыSatisfacción salarial clave permanencia trabajadoresGuillermo MonteroОценок пока нет
- Satisfacción salarial clave permanencia trabajadoresДокумент2 страницыSatisfacción salarial clave permanencia trabajadoresGuillermo MonteroОценок пока нет
- Proyecto Psicologia Clinica Primera EntregaДокумент16 страницProyecto Psicologia Clinica Primera EntregaMaritza mojicaОценок пока нет
- Entrega Final Metodos de AnalisisДокумент27 страницEntrega Final Metodos de AnalisisMaritza yadira Mojica avilaОценок пока нет
- Qué Es y Cómo Se Realiza Un EnsayoДокумент4 страницыQué Es y Cómo Se Realiza Un EnsayoJosé Antonio EstrellaОценок пока нет
- Satisfacción salarial clave permanencia trabajadoresДокумент2 страницыSatisfacción salarial clave permanencia trabajadoresGuillermo MonteroОценок пока нет
- Proyecto Psicologia Clinica Primera EntregaДокумент16 страницProyecto Psicologia Clinica Primera EntregaMaritza mojicaОценок пока нет
- Tercera Entrega MRPДокумент53 страницыTercera Entrega MRPMaritza mojicaОценок пока нет
- Primera Entrega Psicologia ClinicaДокумент11 страницPrimera Entrega Psicologia ClinicaMaritza yadira Mojica avilaОценок пока нет
- AdminverblobawaДокумент142 страницыAdminverblobawaJAIMEОценок пока нет
- Tercera Entrega Psicologia ClinicaДокумент38 страницTercera Entrega Psicologia ClinicaMaritza mojicaОценок пока нет
- Segunda Entrega Psicologia ClinicaДокумент1 страницаSegunda Entrega Psicologia ClinicaMaritza mojicaОценок пока нет
- Cómo prevenir el fracaso escolar de estudiantes de primaria durante el confinamiento por COVID-19 a través de un enfoque conductistaДокумент4 страницыCómo prevenir el fracaso escolar de estudiantes de primaria durante el confinamiento por COVID-19 a través de un enfoque conductistaMaritza yadira Mojica avilaОценок пока нет
- Tercera Entrega Psicologia Social y Comunitaria TURNITINДокумент41 страницаTercera Entrega Psicologia Social y Comunitaria TURNITINMaritza mojicaОценок пока нет
- Marco Teorico Psicologia Social.Документ4 страницыMarco Teorico Psicologia Social.Maritza mojicaОценок пока нет
- Cómo prevenir el fracaso escolar de estudiantes de primaria durante el confinamiento por COVID-19 a través de un enfoque conductistaДокумент4 страницыCómo prevenir el fracaso escolar de estudiantes de primaria durante el confinamiento por COVID-19 a través de un enfoque conductistaMaritza yadira Mojica avilaОценок пока нет
- Primera Entrega ComportamientoДокумент6 страницPrimera Entrega ComportamientoMaritza mojicaОценок пока нет
- Animales de Las Regiones de EcuadorДокумент3 страницыAnimales de Las Regiones de Ecuadorruthsy85Оценок пока нет
- Hoja de Ejercicios 2 RazonamientoДокумент6 страницHoja de Ejercicios 2 RazonamientoIohana Michelle Delgado LemusОценок пока нет
- Alondra CornudaДокумент3 страницыAlondra CornudaGabriela PinzonОценок пока нет
- Tejido OseoДокумент6 страницTejido OseoOswa HumpiriОценок пока нет
- Hemostasia: mecanismos y alteracionesДокумент19 страницHemostasia: mecanismos y alteracionesZetti Lenon Vallejos GomezОценок пока нет
- Anatomía y Biomecánica de La Columna Dorsal y Lumbar o Raquis Dorsal y LumbarДокумент4 страницыAnatomía y Biomecánica de La Columna Dorsal y Lumbar o Raquis Dorsal y LumbarLucho RodriguezОценок пока нет
- Sistema Nervioso de Crustaceos y ArañasДокумент34 страницыSistema Nervioso de Crustaceos y ArañasMairimMikeMtai50% (2)
- CUADERNILLO N°4 CIENCIAS 2°basico JUNIOДокумент13 страницCUADERNILLO N°4 CIENCIAS 2°basico JUNIONICOLEОценок пока нет
- No Te Metas Con Los Gatos Realismo MagicoДокумент3 страницыNo Te Metas Con Los Gatos Realismo MagicoNinia AbacaxiОценок пока нет
- TIBURONESДокумент14 страницTIBURONESAllen RmОценок пока нет
- PRACTICA, CÁLCULO DE DISEÑO y Armado de Redes Tipo CortinaДокумент13 страницPRACTICA, CÁLCULO DE DISEÑO y Armado de Redes Tipo Cortinakhintia mulatoОценок пока нет
- LaVerdaderaHistoria Del HalfMoonДокумент7 страницLaVerdaderaHistoria Del HalfMoonBettaSplendenMx1Оценок пока нет
- Ordenes de Insectos Exposición 1Документ10 страницOrdenes de Insectos Exposición 1Hugo AguilarОценок пока нет
- Anatomía humana: Ejercicio de direcciones muscularesДокумент1 страницаAnatomía humana: Ejercicio de direcciones muscularesVelázquez Pérez Ángela ArisbelОценок пока нет
- Binomio LatinoДокумент29 страницBinomio LatinoAgrnomoОценок пока нет
- Diametro de OvocitosДокумент3 страницыDiametro de OvocitosSergio FerОценок пока нет
- Delfines en el zoológicoДокумент12 страницDelfines en el zoológicogloria yaneth morales castrillonОценок пока нет
- Manual de Oftalmologia General 1 U Chile PDFДокумент125 страницManual de Oftalmologia General 1 U Chile PDFPamela Del Pilar BetancourtОценок пока нет
- Biologia Celular (BL) (BQ)Документ3 страницыBiologia Celular (BL) (BQ)nigax XcОценок пока нет
- Exploración Radiográfica de Columna Lumbar, Sacro y Cóccix.Документ21 страницаExploración Radiográfica de Columna Lumbar, Sacro y Cóccix.Julio Cesar ACОценок пока нет
- Resumen Coprocultivo 4101c3Документ22 страницыResumen Coprocultivo 4101c3Jimmy Moyolema0% (1)
- yZLq - Aparato Circulatorio Animales (Oxford)Документ8 страницyZLq - Aparato Circulatorio Animales (Oxford)Astrid OrtizОценок пока нет
- Procesos de osificación ósea y factores que afectan la densidad mineral óseaДокумент3 страницыProcesos de osificación ósea y factores que afectan la densidad mineral óseaVanessaYepezPerezОценок пока нет
- Músculos cabeza funcionesДокумент14 страницMúsculos cabeza funcionesElena VillanuevaОценок пока нет