Вам также может понравиться
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (120)
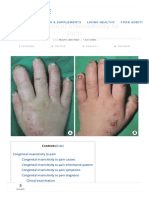
- Achondroplasia Stacy, NikkyДокумент14 страницAchondroplasia Stacy, NikkyStacy PhuongОценок пока нет
- Congenital Hydrocephalus in Clinical Practice EngДокумент7 страницCongenital Hydrocephalus in Clinical Practice Engtugam umarОценок пока нет
- HSC Biology Module 8Документ10 страницHSC Biology Module 8arabellatav23Оценок пока нет
- Movement Disorders in Children: Continuumaudio Interview Available OnlineДокумент22 страницыMovement Disorders in Children: Continuumaudio Interview Available OnlineSnezana MihajlovicОценок пока нет
- Cad Report Human Rights - FinalДокумент35 страницCad Report Human Rights - FinalJohn Paul CairelОценок пока нет
- Genetics and Genetic Counselling: Gloria P. de Leon, RN, RM, ManДокумент64 страницыGenetics and Genetic Counselling: Gloria P. de Leon, RN, RM, ManJoan VillafrancaОценок пока нет
- Congenital Insensitivity To Pain Causes, Symptoms, Diagnosis & TДокумент45 страницCongenital Insensitivity To Pain Causes, Symptoms, Diagnosis & TPan DjeroОценок пока нет
- Intolerancia A La Fructosa Artículo RevisiónДокумент22 страницыIntolerancia A La Fructosa Artículo RevisiónCarol100% (1)
- De de Xuat HV 11 2018-2019Документ14 страницDe de Xuat HV 11 2018-2019Alin NguyenОценок пока нет
- Developmental psychologyEGC 803Документ127 страницDevelopmental psychologyEGC 803Jones DavidОценок пока нет
- Cmca Lec PrelimДокумент22 страницыCmca Lec PrelimAnyrОценок пока нет
- GENETIC COUNSELLING (FARHEEN KHAN MSC 1st Year)Документ8 страницGENETIC COUNSELLING (FARHEEN KHAN MSC 1st Year)Farheen khanОценок пока нет
- 1 Thoracic and Abdominal AneurysmДокумент37 страниц1 Thoracic and Abdominal AneurysmErneslito LucasОценок пока нет
- Chapter23-26 - Defence Against Diseases PDFДокумент10 страницChapter23-26 - Defence Against Diseases PDFBilly LamОценок пока нет
- Pediatrics: by DR - Mohammad Z. Abu Sheikha@Документ98 страницPediatrics: by DR - Mohammad Z. Abu Sheikha@Mohamed AbbasОценок пока нет
- Congenital Fibrosis of The Extra-Ocular Muscles Syndrome Review ArticleДокумент5 страницCongenital Fibrosis of The Extra-Ocular Muscles Syndrome Review Articleanon_703701023Оценок пока нет
- Human Genetic Disorders: and PedigreesДокумент51 страницаHuman Genetic Disorders: and Pedigreescm4Оценок пока нет
- Cystic Fibrosis Fact SheetДокумент6 страницCystic Fibrosis Fact SheetAnggun SafariantiniОценок пока нет
- Etiology of MalocclusionДокумент185 страницEtiology of MalocclusionSumita Jain100% (2)
- BI2332 Concepts of Disease-Congenital Adrenal Hyperplasia WorkshopДокумент7 страницBI2332 Concepts of Disease-Congenital Adrenal Hyperplasia WorkshopFrances EmmanuelОценок пока нет
- Inborn Error of MetabolismДокумент33 страницыInborn Error of MetabolismAlmiraОценок пока нет
- Human Existence and Ethics: PreservationДокумент2 страницыHuman Existence and Ethics: PreservationZabdiel Ann SavellanoОценок пока нет
- Peter Conrad, A Mirage of GenesДокумент14 страницPeter Conrad, A Mirage of GenesKein BécilОценок пока нет
- Brenda Greyling Genetic Counsellor, MSC (Med) National Health Laboratory Service & WitsДокумент4 страницыBrenda Greyling Genetic Counsellor, MSC (Med) National Health Laboratory Service & WitsNeo Mervyn MonahengОценок пока нет
- Biology ProjectДокумент23 страницыBiology ProjectJay Prakash100% (3)
- Contribution of Muscle MRI For Diagnosis of MyopathyДокумент20 страницContribution of Muscle MRI For Diagnosis of MyopathyJose EdgarОценок пока нет
- Genetic Counselling PDFДокумент7 страницGenetic Counselling PDFTanvi100% (1)
- Form B: Non-Invasive InvasiveДокумент12 страницForm B: Non-Invasive InvasiveSrikanth KanagasabapathyОценок пока нет
- Startup Inspiration Black by SlidesgoДокумент7 страницStartup Inspiration Black by SlidesgoKrystel LedesmaОценок пока нет
- McAllister Genetics ProjectДокумент4 страницыMcAllister Genetics ProjectTysonОценок пока нет