Вам также может понравиться
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- SongbookДокумент123 страницыSongbookRtoes100% (3)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Shakti - The Spiritual Science of DNA by E. J. GoldДокумент48 страницShakti - The Spiritual Science of DNA by E. J. GoldstimoceiverОценок пока нет
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Roy Adaptation Model For Nursing Had ItДокумент4 страницыThe Roy Adaptation Model For Nursing Had ItRazaCreciaLastrillaMenesesОценок пока нет
- Understanding Culture, Society and Politics: Fourth QuarterДокумент21 страницаUnderstanding Culture, Society and Politics: Fourth QuarterGordon Gorospe100% (1)
- A Natural History of Rape - Biological Bases of Sexual Coercion: Randy Thornhill & Craig T. PalmerДокумент270 страницA Natural History of Rape - Biological Bases of Sexual Coercion: Randy Thornhill & Craig T. PalmerDeep Narayan Chatterjee100% (1)
- Ecosystems at Risk UnitДокумент136 страницEcosystems at Risk UnitJess IsaacsОценок пока нет
- Bio 345 Evolution Module 1 & 2 Exam 1 Review GuideДокумент17 страницBio 345 Evolution Module 1 & 2 Exam 1 Review GuideTiffanie100% (1)
- Cal Spring 13sylaДокумент2 страницыCal Spring 13sylaRtoesОценок пока нет
- Lang Policy in SAДокумент17 страницLang Policy in SARtoesОценок пока нет
- Final PPG NursingДокумент4 страницыFinal PPG NursingRtoesОценок пока нет
- Culinary Arts Program /CALENDAR / Homework Assignment Subject To ChangeДокумент2 страницыCulinary Arts Program /CALENDAR / Homework Assignment Subject To ChangeRtoesОценок пока нет
- The Evolution of Language: Michael C. CorballisДокумент25 страницThe Evolution of Language: Michael C. CorballisRtoesОценок пока нет
- LSSyllabusДокумент3 страницыLSSyllabusRtoesОценок пока нет
- Week 7Документ4 страницыWeek 7Rtoes0% (1)
- Languages Are Vanishing: So What?: Story in The News Magazine The EconomistДокумент8 страницLanguages Are Vanishing: So What?: Story in The News Magazine The EconomistRtoesОценок пока нет
- Zuckerman 2006Документ16 страницZuckerman 2006RtoesОценок пока нет
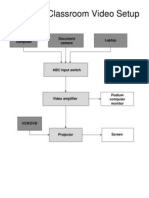
- Standard Classroom VideoДокумент1 страницаStandard Classroom VideoRtoesОценок пока нет
- NIH Public Access: Author ManuscriptДокумент24 страницыNIH Public Access: Author ManuscriptRtoesОценок пока нет
- Neutrality of International Languages: Haitao LiuДокумент28 страницNeutrality of International Languages: Haitao LiuRtoesОценок пока нет
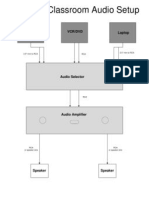
- Standard Classroom AudioДокумент1 страницаStandard Classroom AudioRtoesОценок пока нет
- Lewis&Elman (2001)Документ12 страницLewis&Elman (2001)RtoesОценок пока нет
- Unit 1 ReviewДокумент18 страницUnit 1 ReviewZeeОценок пока нет
- Worksheet Q3 Week 6Документ2 страницыWorksheet Q3 Week 6Jaybie TejadaОценок пока нет
- Parasites Immunity and PathologyДокумент389 страницParasites Immunity and PathologyCarlos Bell Castro Tamayo100% (1)
- Grade10 - Las4 - Final VersionДокумент4 страницыGrade10 - Las4 - Final VersionLester Keno LejanoОценок пока нет
- Handbook For The New Paradigm - 02Документ59 страницHandbook For The New Paradigm - 02Τζένη ΔασκαλοπούλουОценок пока нет
- Q3 Science LLCДокумент20 страницQ3 Science LLCRoger BaliliОценок пока нет
- 5399 5668 02Документ19 страниц5399 5668 02Ashok KarriОценок пока нет
- Commoner's Law of EcologyДокумент8 страницCommoner's Law of EcologyPau Figuerres0% (1)
- LLED 360 - Life Science 11 Unit Plan - EvolutionДокумент24 страницыLLED 360 - Life Science 11 Unit Plan - EvolutionamandaОценок пока нет
- Science Year 5: Work Sample PortfolioДокумент30 страницScience Year 5: Work Sample Portfolioallan.manaloto23Оценок пока нет
- Charles DarwinДокумент11 страницCharles DarwinJeon TaeTaeОценок пока нет
- Animal Adaptations: Lesson Plans and Activities For Your Classroom and While Visiting The ZooДокумент6 страницAnimal Adaptations: Lesson Plans and Activities For Your Classroom and While Visiting The ZooLove Mie MoreОценок пока нет
- Johan Olsen-The Many Faces of EuropeanizationДокумент36 страницJohan Olsen-The Many Faces of EuropeanizationbbmikОценок пока нет
- Adapting To SurviveДокумент2 страницыAdapting To SurviveFaiez Ali33% (3)
- Handout For Natural Selection WebquestДокумент6 страницHandout For Natural Selection WebquestLisa Lopez ArguetaОценок пока нет
- Learning Progressions Group Assignment Itl 518Документ18 страницLearning Progressions Group Assignment Itl 518api-421764959Оценок пока нет
- Science Reviewer For Gr.3Документ43 страницыScience Reviewer For Gr.3ARISTHEA C. LABRADORОценок пока нет
- Week 1 Lecture 1 BIOCHEMISTRYДокумент96 страницWeek 1 Lecture 1 BIOCHEMISTRYaira sharidaОценок пока нет
- 涛博GRE结班测试 笔记版Документ41 страница涛博GRE结班测试 笔记版geyirucОценок пока нет
- A Hoot in The DarkДокумент22 страницыA Hoot in The DarkDustin Greenwalt100% (1)
- Exercise 7 - Evolution and SystematicsДокумент8 страницExercise 7 - Evolution and SystematicsPaulo SantosОценок пока нет
- NURS800 Myra Levine Conservation Model-1Документ12 страницNURS800 Myra Levine Conservation Model-1risqi wahyu100% (1)
- Aquatic Adaptations - Poonam SinghДокумент46 страницAquatic Adaptations - Poonam Singhaksahu01234Оценок пока нет
- Behavioral Plasticity. An Interaction Between Evolution and ExperienceДокумент13 страницBehavioral Plasticity. An Interaction Between Evolution and ExperiencevmsolartecОценок пока нет