Вам также может понравиться
- Biology of Implant OsseointegrationДокумент11 страницBiology of Implant OsseointegrationAmIrОценок пока нет
- Ehab ThesisДокумент133 страницыEhab Thesisehabmohamedzayed100% (2)
- BIO 192 Lab Report #1 BivalveДокумент8 страницBIO 192 Lab Report #1 BivalveBen KillamОценок пока нет
- Biodiversity & StabilityДокумент76 страницBiodiversity & StabilityMAXINE CLAIRE CUTINGОценок пока нет
- PNAS 1997 Yu 9226 31Документ6 страницPNAS 1997 Yu 9226 31Dani Pazmiño BonillaОценок пока нет
- Comparative Population Genomics of Maize Domestication and Improvement Hufford Et Al. 2012Документ6 страницComparative Population Genomics of Maize Domestication and Improvement Hufford Et Al. 2012Cindi ContrerasОценок пока нет
- Megabase Level Sequencing Reveals Contrasted Organization and Evolution Patterns of The Wheat Gene and Transposable Element SpacesДокумент16 страницMegabase Level Sequencing Reveals Contrasted Organization and Evolution Patterns of The Wheat Gene and Transposable Element Spaces10sgОценок пока нет
- Broad-Scale Recombination Pattern in The Primitive Bird Rhea Americana (Ratites, Palaeognathae)Документ13 страницBroad-Scale Recombination Pattern in The Primitive Bird Rhea Americana (Ratites, Palaeognathae)lutfiamegawatiОценок пока нет
- tmpAC TMPДокумент12 страницtmpAC TMPFrontiersОценок пока нет
- Sanmiguel, 1998Документ8 страницSanmiguel, 1998Chie SakuraОценок пока нет
- Genetic Divergence Among Maize (Zea Mays L.) Inbred Lines Using Morphometric MarkersДокумент15 страницGenetic Divergence Among Maize (Zea Mays L.) Inbred Lines Using Morphometric MarkersIJAR JOURNALОценок пока нет
- AndyGWAS-RevisedManuscript FileДокумент26 страницAndyGWAS-RevisedManuscript FileZeleke WondimuОценок пока нет
- Ijms 21 01344 v2Документ17 страницIjms 21 01344 v2Diane Fernandez BenesioОценок пока нет
- Phenotypic Molecular and Biochemical Evaluation ofДокумент11 страницPhenotypic Molecular and Biochemical Evaluation ofAmina BelguendouzОценок пока нет
- Estimating Genome Conservation Between Crop and Model Legume SpeciesДокумент6 страницEstimating Genome Conservation Between Crop and Model Legume SpeciesAnny LovibОценок пока нет
- 1989 TagДокумент6 страниц1989 TaglacisagОценок пока нет
- The Effect of Storage Environment On Seed Survival and The Accumulation of Chromosomal Aberrations in Pea Landraces and Cultivars (Pisum Sativum L.)Документ10 страницThe Effect of Storage Environment On Seed Survival and The Accumulation of Chromosomal Aberrations in Pea Landraces and Cultivars (Pisum Sativum L.)Ariana ChimiОценок пока нет
- One Thousand Plant Transcriptomes and Phylogenomics, 2019Документ20 страницOne Thousand Plant Transcriptomes and Phylogenomics, 2019fosudavid.jfОценок пока нет
- Co-So-Di-Truyen-Chon-Giong-Cay-Trong - Agronomy-10-00180 - (Cuuduongthancong - Com)Документ15 страницCo-So-Di-Truyen-Chon-Giong-Cay-Trong - Agronomy-10-00180 - (Cuuduongthancong - Com)Hai HeoОценок пока нет
- 05 MK Biswas Et Al in Exploaitation of GeneticДокумент16 страниц05 MK Biswas Et Al in Exploaitation of GeneticDaniella MotusicОценок пока нет
- 00150526Документ11 страниц00150526Ines Jlassi Ep MabroukОценок пока нет
- Plant Genome Projects Focus on Model OrganismsДокумент4 страницыPlant Genome Projects Focus on Model Organismsmanoj_rkl_07100% (1)
- BCAT-based Marker For Marker-Assisted Selection in Vietnam Cucumber BreedingДокумент5 страницBCAT-based Marker For Marker-Assisted Selection in Vietnam Cucumber BreedingShailendra RajanОценок пока нет
- PNAS 1998 Martienssen 2021 6Документ6 страницPNAS 1998 Martienssen 2021 6Aseliajulia JuliaОценок пока нет
- Thermo-Sensitive Wild Aborted Cytoplasmic Genic Male Sterility in Rice (Oryza Sativa L.) Having Majority Panicles Completely ExsertedДокумент11 страницThermo-Sensitive Wild Aborted Cytoplasmic Genic Male Sterility in Rice (Oryza Sativa L.) Having Majority Panicles Completely ExsertedPremier PublishersОценок пока нет
- 1 s2.0 S1674205218303642 MainДокумент3 страницы1 s2.0 S1674205218303642 MainRodrigo RuizОценок пока нет
- ArtikelДокумент12 страницArtikelRahmawati LailiОценок пока нет
- Plant Functional Genomics 99Документ4 страницыPlant Functional Genomics 99Vanessa GroberОценок пока нет
- BMC Evolutionary BiologyДокумент10 страницBMC Evolutionary Biology10sgОценок пока нет
- Genetic Diversity of Recombinant Inbred Lines of Phaseolus Vulgaris LДокумент9 страницGenetic Diversity of Recombinant Inbred Lines of Phaseolus Vulgaris LfranciscoОценок пока нет
- Ijms 21113778Документ20 страницIjms 21113778Alejandro GutiérrezОценок пока нет
- Badouin 2017 Nat The Sunflower Geno Prov inДокумент14 страницBadouin 2017 Nat The Sunflower Geno Prov inAJMRОценок пока нет
- Morell Ibarra Et Al. Crop Genomics Advances and ApplicationsДокумент12 страницMorell Ibarra Et Al. Crop Genomics Advances and ApplicationswillnotburnОценок пока нет
- Complete Nucleotide Sequence of The Sugarcane (Four Monocot Chloroplast GenomesДокумент7 страницComplete Nucleotide Sequence of The Sugarcane (Four Monocot Chloroplast GenomesmemorieleОценок пока нет
- Pissard Et Al 2007 Oca DiversidadДокумент15 страницPissard Et Al 2007 Oca DiversidadBlaxez YTОценок пока нет
- SSR Potato Genetic DiversityДокумент9 страницSSR Potato Genetic DiversityM. Rehman Gul KhanОценок пока нет
- Diversity Assessment of Yield Yield Contributing Traits and Earliness of Advanced T-Aman Rice Oryza Sativa L. LinesДокумент11 страницDiversity Assessment of Yield Yield Contributing Traits and Earliness of Advanced T-Aman Rice Oryza Sativa L. Linesনাজমুল হক শাহিনОценок пока нет
- JSS 919 PublicadoДокумент9 страницJSS 919 PublicadocAROLINAОценок пока нет
- Bainard Et Al 2020. Genome Size and Endopolyploidy Evolution Across The Moss PhylogenyДокумент13 страницBainard Et Al 2020. Genome Size and Endopolyploidy Evolution Across The Moss PhylogenylaurachalupОценок пока нет
- Cytotaxonomy and Karyotype EvolutionДокумент12 страницCytotaxonomy and Karyotype EvolutionJomaira MedinaОценок пока нет
- Practical Report AboutImitation Ratio Phenotype - Yolanda AnggraeniДокумент12 страницPractical Report AboutImitation Ratio Phenotype - Yolanda Anggraeniyolanda anggraeniОценок пока нет
- University of Agricultural Sciences, Dharwad Department of Genetics and Plant BreedingДокумент8 страницUniversity of Agricultural Sciences, Dharwad Department of Genetics and Plant Breedingkavyashree g mОценок пока нет
- Assessment of Genetic Variability and Heritability in Segregating PopuationsДокумент4 страницыAssessment of Genetic Variability and Heritability in Segregating PopuationsTJPRC PublicationsОценок пока нет
- Study of Genetic Variability and Character Association For Yield and Yield Related Traits in f3 Generation of Blackgram (Vignamungo (L.) Hepper)Документ13 страницStudy of Genetic Variability and Character Association For Yield and Yield Related Traits in f3 Generation of Blackgram (Vignamungo (L.) Hepper)Mamta AgarwalОценок пока нет
- Grain Protein Content Variation and Its Association Analysis in BarleyДокумент11 страницGrain Protein Content Variation and Its Association Analysis in BarleySyrine SomraniОценок пока нет
- SSRN Id4390423Документ14 страницSSRN Id4390423Chie SakuraОценок пока нет
- Ground Nut Research in Odisha, A Case Study and AnalysisДокумент16 страницGround Nut Research in Odisha, A Case Study and AnalysisTJPRC PublicationsОценок пока нет
- Simmonds 2014Документ13 страницSimmonds 2014Victor BonillaОценок пока нет
- Frontiers TemplateДокумент4 страницыFrontiers TemplateRichard Estrada C.Оценок пока нет
- Correa Et Al., 2014 TAG 10.1007 - S00122-014-2286-YДокумент22 страницыCorrea Et Al., 2014 TAG 10.1007 - S00122-014-2286-Yf.oyarce.caroОценок пока нет
- RM - 171Документ12 страницRM - 171Mehvish ChОценок пока нет
- Assessment of genetic diversity and relationship of coastal salt tolerant rice accessions of KeralaДокумент8 страницAssessment of genetic diversity and relationship of coastal salt tolerant rice accessions of KeralaWence G. HerreraОценок пока нет
- Genetic Variability in Elite Barley Genotypes Based On The Agro-Morphological Characteristics Evaluated Under Irrigated SystemДокумент12 страницGenetic Variability in Elite Barley Genotypes Based On The Agro-Morphological Characteristics Evaluated Under Irrigated SystemGiselle RodolfoОценок пока нет
- Studies On Variability and Heritability in RiceДокумент2 страницыStudies On Variability and Heritability in RiceDARSHANA AJITHОценок пока нет
- Rajalakshmi Engineering College Plant Biotechnology NotesДокумент51 страницаRajalakshmi Engineering College Plant Biotechnology Noteskrishradhan1100% (1)
- 40 Ijasrapr201740Документ8 страниц40 Ijasrapr201740TJPRC PublicationsОценок пока нет
- Rayburn Genome SizeДокумент6 страницRayburn Genome SizeSebastian DavisОценок пока нет
- Thesis - Suhu Tinggi - NatawijayaДокумент180 страницThesis - Suhu Tinggi - NatawijayaUsamah JaisyurahmanОценок пока нет
- 2015 Article 1815Документ15 страниц2015 Article 1815Houda LaatabiОценок пока нет
- Molecular Versus Morphological Markers To Describe Variability in Sugar Cane (Saccharum Officinarum) For Germplasm Management and ConservationДокумент21 страницаMolecular Versus Morphological Markers To Describe Variability in Sugar Cane (Saccharum Officinarum) For Germplasm Management and ConservationBlaxez YTОценок пока нет
- Phylogenetic Studies in MycorrhizasДокумент17 страницPhylogenetic Studies in Mycorrhizasrahulsinghrs100% (1)
- Transferring Hessian Fly Resistance from Aegilops geniculata to Durum WheatДокумент6 страницTransferring Hessian Fly Resistance from Aegilops geniculata to Durum WheatMuhammad Luqman KuswardanaОценок пока нет
- HEC Voucher NewДокумент1 страницаHEC Voucher NewAyyan AyyanОценок пока нет
- PESTICIDE REGISTERED LIST - Form 1 PDFДокумент24 страницыPESTICIDE REGISTERED LIST - Form 1 PDFmuzamil rajpoot67% (3)
- Maize (Zea Mays L.) Genome Size Indicated by 180-bp Knob Abundance Is Associated With Flowering TimeДокумент9 страницMaize (Zea Mays L.) Genome Size Indicated by 180-bp Knob Abundance Is Associated With Flowering TimeAyyan AyyanОценок пока нет
- Maize (Zea Mays L.) Genome Size Indicated by 180-bp Knob Abundance Is Associated With Flowering TimeДокумент9 страницMaize (Zea Mays L.) Genome Size Indicated by 180-bp Knob Abundance Is Associated With Flowering TimeAyyan AyyanОценок пока нет
- Adolescent Reproductive and Sexual HealthДокумент42 страницыAdolescent Reproductive and Sexual HealthMuhammad Abbas WaliОценок пока нет
- LWT - Food Science and TechnologyДокумент8 страницLWT - Food Science and TechnologyAzb 711Оценок пока нет
- Brigitte King - InformativeДокумент2 страницыBrigitte King - Informativeapi-262025468Оценок пока нет
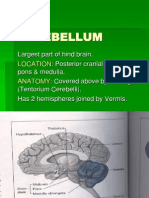
- Physiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamДокумент61 страницаPhysiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamZaigham HammadОценок пока нет
- Cellular RespirationДокумент20 страницCellular RespirationAlessiaОценок пока нет
- (PDF) Root Growth of Phalsa (Grewia Asiatica L.) As Affected by Type of CuttДокумент5 страниц(PDF) Root Growth of Phalsa (Grewia Asiatica L.) As Affected by Type of CuttAliОценок пока нет
- Protected Area Expansion Strategy For CapeNature 2010 29 SeptemberДокумент70 страницProtected Area Expansion Strategy For CapeNature 2010 29 SeptembertableviewОценок пока нет
- C Specialized GeneralBiology1 Sem1 Clas3 CellsinPlantand-AnimalTissue v2Документ14 страницC Specialized GeneralBiology1 Sem1 Clas3 CellsinPlantand-AnimalTissue v2ダギア メイ アニアОценок пока нет
- Psychiatric Mental Health Nursing Concepts of Care in Evidence Based Practice 8th Edition Townsend Test BankДокумент36 страницPsychiatric Mental Health Nursing Concepts of Care in Evidence Based Practice 8th Edition Townsend Test Bankquemefuloathableilljzf100% (26)
- Met. Secundarios Cáscara SagradaДокумент9 страницMet. Secundarios Cáscara SagradaDalíAsesoríasОценок пока нет
- Dna Finger PrintingДокумент23 страницыDna Finger PrintingSanjay KumarОценок пока нет
- 2 Part 1Документ99 страниц2 Part 1m_luchianОценок пока нет
- Plant Transport - IGCSE Biology Notes (2020)Документ6 страницPlant Transport - IGCSE Biology Notes (2020)Zhi En LeeОценок пока нет
- Paul Atriedes (PL 6/181pp) : Bene Gesserit Training (Container, Active 11)Документ2 страницыPaul Atriedes (PL 6/181pp) : Bene Gesserit Training (Container, Active 11)dfgdfОценок пока нет
- Lesson Plan No 5Документ6 страницLesson Plan No 5hafeez ahmedОценок пока нет
- Bioinformatic Analysis of HBBДокумент1 страницаBioinformatic Analysis of HBBfrank2689Оценок пока нет
- Life in The Deserts Worksheet 2Документ2 страницыLife in The Deserts Worksheet 2Uma TiwariОценок пока нет
- 8 Cell - The Unit of Life-NotesДокумент6 страниц8 Cell - The Unit of Life-NotesBhavanya RavichandrenОценок пока нет
- Follicular Dynamics in Bovine and Ovine 1Документ19 страницFollicular Dynamics in Bovine and Ovine 1israr yousafОценок пока нет
- 10th Standard Tamilnadu State Board Science (English Medium)Документ283 страницы10th Standard Tamilnadu State Board Science (English Medium)Karthick NОценок пока нет
- Jurnal Perkecambahan Biji PDFДокумент14 страницJurnal Perkecambahan Biji PDFKinad Danik50% (2)
- Komiyama Et Al., (2008) Allometry, Biomass, and Productivity of Mangrove Forests A ReviewДокумент11 страницKomiyama Et Al., (2008) Allometry, Biomass, and Productivity of Mangrove Forests A ReviewVandhi Amali100% (1)
- MYCOVIROДокумент11 страницMYCOVIROPau SorianoОценок пока нет
- 2nd Periodic Test (Science)Документ1 страница2nd Periodic Test (Science)mosarbas0950% (2)
- Gene TherapyДокумент1 страницаGene TherapyJenevieve B. CañeteОценок пока нет
- Chapter 5Документ14 страницChapter 5Caitlin G.Оценок пока нет
- Molecular Cloning Technical GuideДокумент40 страницMolecular Cloning Technical GuideRОценок пока нет