Вам также может понравиться
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- 6th GATEДокумент33 страницы6th GATESamejiel Aseviel LajesielОценок пока нет
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Jt20 ManualДокумент201 страницаJt20 Manualweider11Оценок пока нет
- Fisiologia de KatzДокумент663 страницыFisiologia de KatzOscar Gascon100% (1)
- Trend Graphs - Sample AnswerДокумент4 страницыTrend Graphs - Sample AnswerannieannsОценок пока нет
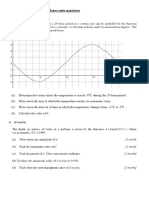
- Sine and Cosine Exam QuestionsДокумент8 страницSine and Cosine Exam QuestionsGamer Shabs100% (1)
- Express VPN Activation CodeДокумент5 страницExpress VPN Activation CodeButler49JuulОценок пока нет
- Facebook Facing Off Againt TencentДокумент6 страницFacebook Facing Off Againt TencentWai Shan Lee0% (1)
- The Unseelie Prince Maze of Shadows Book 1 by Kathryn AnnДокумент267 страницThe Unseelie Prince Maze of Shadows Book 1 by Kathryn Annanissa Hri50% (2)
- Method Statement Pressure TestingДокумент15 страницMethod Statement Pressure TestingAkmaldeen AhamedОценок пока нет
- Securing Obligations Through Pledge and MortgageДокумент4 страницыSecuring Obligations Through Pledge and MortgagePrincessAngelaDeLeon100% (1)
- NYU Stern Evaluation NewsletterДокумент25 страницNYU Stern Evaluation NewsletterCanadianValueОценок пока нет
- Nistha Tamrakar Chicago Newa VIIДокумент2 страницыNistha Tamrakar Chicago Newa VIIKeshar Man Tamrakar (केशरमान ताम्राकार )Оценок пока нет
- PronPack 5 Sample MaterialДокумент13 страницPronPack 5 Sample MaterialAlice FewingsОценок пока нет
- Pakistan Affairs Current Affairs 2016 MCQSДокумент3 страницыPakistan Affairs Current Affairs 2016 MCQSMuhammad MudassarОценок пока нет
- Reg OPSДокумент26 страницReg OPSAlexandru RusuОценок пока нет
- Compliance Risks and Controls: Speaker: Dott. Fabio AccardiДокумент14 страницCompliance Risks and Controls: Speaker: Dott. Fabio AccardiJefferson WidodoОценок пока нет
- Nigerian Romance ScamДокумент10 страницNigerian Romance ScamAnonymous Pb39klJОценок пока нет
- Pakage PAU (Fruits)Документ70 страницPakage PAU (Fruits)Gary Bhullar57% (7)
- 09 Lift Cylinder Drift (Bulldozer) - CheckДокумент2 страницы09 Lift Cylinder Drift (Bulldozer) - CheckFredy Manrique AstoОценок пока нет
- Froyen06-The Keynesian System I - The Role of Aggregate DemandДокумент40 страницFroyen06-The Keynesian System I - The Role of Aggregate DemandUditi BiswasОценок пока нет
- 10 Compactness in Function Spaces: Ascoli-Arzel A TheoremДокумент5 страниц10 Compactness in Function Spaces: Ascoli-Arzel A TheoremronalduckОценок пока нет
- EHT For Athletes For Brain HealthДокумент2 страницыEHT For Athletes For Brain HealthTracey Jorg RollisonОценок пока нет
- Texas LS Notes 19-20Документ2 страницыTexas LS Notes 19-20Jesus del CampoОценок пока нет
- Dxgbvi Abdor Rahim OsmanmrДокумент1 страницаDxgbvi Abdor Rahim OsmanmrSakhipur TravelsОценок пока нет
- Unit 4 AE Scan OCRed PDFДокумент44 страницыUnit 4 AE Scan OCRed PDFZia AhmedОценок пока нет
- Linear Circuit Analysis (ELEN-1100) : Lecture # 13: More On Mesh Current AnalysisДокумент11 страницLinear Circuit Analysis (ELEN-1100) : Lecture # 13: More On Mesh Current AnalysisPhD EEОценок пока нет
- Electronic Throttle ControlДокумент67 страницElectronic Throttle Controlmkisa70100% (1)
- Course Code: Hrm353 L1Документ26 страницCourse Code: Hrm353 L1Jaskiran KaurОценок пока нет
- Viviana Rodriguez: Education The University of Texas at El Paso (UTEP)Документ1 страницаViviana Rodriguez: Education The University of Texas at El Paso (UTEP)api-340240168Оценок пока нет
- Reasons for Conducting Qualitative ResearchДокумент12 страницReasons for Conducting Qualitative ResearchMa. Rhona Faye MedesОценок пока нет