Академический Документы
Профессиональный Документы
Культура Документы
Desarrollo Atm y SNC
Загружено:
mariellaevaОригинальное название
Авторское право
Доступные форматы
Поделиться этим документом
Поделиться или встроить документ
Этот документ был вам полезен?
Это неприемлемый материал?
Пожаловаться на этот документАвторское право:
Доступные форматы
Desarrollo Atm y SNC
Загружено:
mariellaevaАвторское право:
Доступные форматы
DESARROLLO DE ATM Y MADURACION DEL SISTEMA NERVIOSO
INTEGRANTES ESCALANTE CANDIA, ELIZABETH RAMOS LEON, MARIELLA SALAS VELASQUEZ, RAMON SANCHEZ NAVARRO, TANIA
CURSO: CRECIMIENTO Y DESARROLLO ESPECIALIDAD: ORTODONCIA Y ORTOPEDIA MAXILAR PROFESOR: DR. JUSTINIANO SOTOMAYOR
INTRODUCCIN
El siguiente trabajo de los Residentes de primer ao Ortodoncia de la Universidad Inca Garcilazo de la Vega
de la especialidad de tiene como Objetivo
comprender la importancia de 2 fundamentales estructuras: la articulacin temporomandibular y el sistema nervioso.
Debemos de recordar acerca periodonto de insercin, que los componentes que lo conforman son las siguientes estructuras: el cemento, el ligamento periodontal y el Hueso alveolar. Estas tres estructuras conforman un unidad funcional y tienen el mismo origen embriolgico y se forman al mismo tiempo que se forma la raz del diente.
El cemento es el tejido conectivo derivado de la capa celular ectomesenquimtica del folculo dentario que rodea al germen dentario. Desde el punto de vista estructural, el cemento es comparable con el hueso ya que su dureza y composicin qumica son prcticamente similares; adems ambos crecen por aposicin, poseen laminillas, y cuando el cemento presenta clulas estas se alojan en lagunas, tal como lo hacen los osteocitos.
I.
MARCO TEORICO
A)
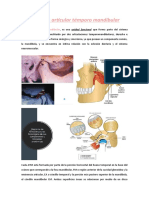
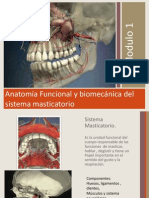
ARTICULACION TEMPOROMANDIBULAR
1. GENERALIDADES
El hueso temporal se relaciona con los huesos del crneo (mediante sinartrosis) por un lado y con el cndilo de Ia mandbula por el otro, conformando con este ltimo una articulacin del tipo de las diartrosis .Por ello, se considera ms apropiado denominar esta conexin del crneo y mandbula, como Complejo Articular temporomandibular (CATM). Cada CATM, est formada, a su vez, por dos articulaciones: una temporodiscal y la otra condIea o discocondilar. El complejo articular temporomandibular (CATM) comprende un conjunto de estructuras anatmicas que, asociadas a grupos musculares permiten la realizacin de los movimientos mandibulares. Desde el punto de vista funcional, el CATM se clasifica como una diartrosis bicondlea, ya que articula dos huesos cuyas superficies convexas se encuentran limitando una cavidad, que contiene un disco articular (como medio de adaptacin) y que est lubricada por el fluido sinovial. Los componentes seos que participan en su constitucin son el cndilo de la mandbula y la eminencia articular del temporal con su fosa mandibular (anteriormente denominada porcin anterior de la cavidad glenoidea) rodeados por una cpsula que protege la articulacin, la cual est reforzada por ligamentos principales y accesorios El CATM es una de las articulaciones ms importantes del organismo, siendo la nica articulacin del cuerpo humano que se caracteriza por trabajar sinrgicamente con la del lado opuesto de forma sincrnica, pudiendo hacerlo de modo independiente si es necesario. El CATM se encuentra ntimamente relacionado con Ia oclusin dentaria y el sistema neuromuscuIar. Por su compleja
dinmica articular, cualquier trastomo funcional o patolgico que asiente en alguno de sus componentes, afectar el normal funcionamiento de todo el sistema.
2. ESTRUCTURA HISTOLOGICA
2.1 Superficies articulares
Estn constituidas por una superficie inferior, el cndilo mandibular, y otra superior, el cndilo del temporal (o raz transversa del cigoma) y Ia cavidad glenoidea, pertenecientes ambas al hueso temporal. La cavidad glenoidea est dividida en dos partes por la cisura de Glaser y, slo la regin anterior es Ia articular, actualmente denominada fosa mandibular (FM). Las reas o superficies articulares destinadas a soportar o resistir las fuerzas mecnicas que se originan durante los movimientos mandibulares, se denominan funcionales. Estas
superficies funcionales estn recubiertas por un tejido conectivo fibroso de mayor espesor, Iocalizado por un lado en la vertiente posterior del cndilo temporal, donde alcanza un grosor de 0,50 mm y a nivel de la carilla articular del cndilo mandibular donde presenta un espesor de 2 mm. Su funcin consiste en amortiguar las presiones y distribuirlas sobre las superficies seas articulares. Las fibras de colgeno (tipo l) superficiales se disrribuyen de forma paralela a las superficies Iibres, mientras que las fibras profundas lo hacen en sentido perpendicular Desde el punto de vista anatmico, el cndilo mandibular es una eminencia elipsoidea, cuyo eje mayor est orientado en sentido oblicuo hacia atrs y adentro. Est unido a la rama mandibular por un segmento estrecho, el cuello del cndilo, que es ms fino en su parte anterointema donde se inserta el msculo pterigoideo externo o lateral. Desde el punto de vista histolgico las superficies articulares estn revestidas, como se ha indicado antes, por una zona de tejido conectivo fibroso, por debajo del cual existe una zona proliferativa muy delgada. Esta capa en el CATM adulto es la que suministra los fibroblastos para renovar el tejido fibroso articular. Subyacente a esta zona proliferativa se observan sucesivamente una zona de fibrocartlago y otra zona muy delgada de cartlago calcificado, tras Ia cual se encuentra el tejido seo subarticular, tanto a nivel mandibular, como temporal. Durante el desarrollo pre y postnatal, eI rea proliferativa de clulas indiferenciadas da tambin origen a los condrocitos subyacentes.
2.2 Disco articular
Representa el medio de adaptacin que tiene por funcin establecer la armona entre las dos superficies articullaresconvexas. Morfolgicamente el disco presenta dos caras, dos bordes y dos extremidades. La cara anterosuperior es cncava por delante (enfrentada a la eminencia temporal) y su parte posterior es convexa (enfrentada a la fosa mandibular). La cara posteroinferior es cncava y cubre al cndilo mandibular por completo. EI borde anterior se contina con el msculo
pterigoideo extemo y recibe fibras de la cpsula articular. El borde posterior y la extremidad externa son ms gruesos. El disco se divide a este nivel en dos lminas proyectando algunas fibras hacia la zona posterior y dirigiendo otras hacia el cuello del cndilo, donde se unen al periostio. Como consecuencia de esta disposicin el disco acompaa al cndilo en todos los movimientos. En la periferia, el disco se conecta con el tejido que forma la cpsula articular y divide a Ia articulacin en dos cavidades sinoviales, supra e infradiscal, ambas con una cinemtica diferente. El disco es delgado en el tercio anterior (I,5 a 2 mm de espesor) y engrosado en los bordes perifricos (2,5 a 3 mm de grosor). La regin ms delgada del disco es la zona central (1 mm), que est compuesta por una densa trama de fibras colgenas, que se ordenan en forma paralela a la superficie articular, junto a la cual existen escasos fibroblastos y ocasionales fibras elsticas. este nivel no se observan vasos sanguneos ni nervios. Los componentes de la matriz amorfa son los que le confieren al disco la capacidad e soportar las fuerzas compresivas, por las propiedades hidroflicas de los proteoglicanos del tipo del condroitn sulfato y dermatn sulfato. En la regin posterior el disco, como indicamos anteriormente, se hace bilaminar y est compuesto por dos fascculos. El fascculo posterosuperior ms desarrollado contiene fibras colgenas, elsticas y algunas fibras reticulares. Por el contrario la lmina posteroinferior del disco, que se une al cndilo es, en cambio, inelstica y avascular. Entre ambos fascculos queda una zona de tejido conectivo laxo, con abundantes vasos sanguneos y nervios. El disco y el cndilo forman una especie de unidad estructural y funcional, ntimamente relacionada con la superficie temporal mediante los ligamentos y msculos asociados. EI borde anterior del disco se halla unido a la fascia y al tendn del msculo pterigoideo lateral. El disco se contina en la periferia con el tejido capsular El disco es flexible y de gran adaptabilidad a los cambios depresibles que experimenta durante el normal funcionamiento. Sin embargo, cuando se producen fuerzas destructoras pequeas y repetidas en el tiempo o cambios estructurales articulares, Ia morfologa del disco puede alterarse irreversiblemente como, por ejemplo, en los disturbios funcionales de traslacin del cndilo mandibular en Ia
apertura bucal. Esta alteracin suele estar presente en casi todas las disfunciones articulares
2.3 Ligamentos y cpsula
Los ligamentos, son estructuras que unen los huesos articulares y que estn constituidas por densos haces de fibras colgenas que se disponen direccionadas en paralelo para soportar mejor las cargas. El CATM tiene ligamentos principales o directos, que intervienen en Ia funcin de Ia misma articulacin y ligamentos de accin indirecta o accesorios, que por sus inserciones restringen en parte la proyeccin anterior de la mandbula. limitando los movimientos condilares. Los ligamentos principales son: l) ligamento capsular, 2) ligamentos colaterales, 3) ligamento temporomandibular y 4) ligamento temporodiscal. Entre los accesorios hay que mencionar: I) el ligamento ptengomandibular, 2) el ligamento esfenomandibular y 3) el ligamento estilomandibular. El ligamento capsular o cpsula, se une por arriba al hueso temporal y por debajo al cndilo, protegiendo de esta manera Ya articulacin. Entre sus funciones, adems de envolver la articulacin, retiene el lquido sinovial, y opone resistencia a cualquier fuerza medial, lateral o vertical inferior, que tienda a separar o luxar las superficies articulares. Desde el punto de vista histolgico, Ia cpsula posee dos capas, una externa fibrosa y una intema muy delgada o membrana sinovial. La cpsula tiene por funcin evitar los movimientos exagerados del cndilo y permitir el desplazamiento del mismo. Hacia fuera, la cpsula se engrosa para formar el ligamento temporomandibular, el cual limita los movimientos mandibulares y se opone a la luxacin durante su actividad funcional. El ligamento temporomandibular es el ms importante de los ligamentos del CATM y consiste en un engrosamiento de la cara lateral de la cpsula. Por su estructura colgena y por Ia presencia ocasional de fibras elsticas, el ligamento es inextensible pero flexible. Refuerza al ligamento capsular, y protege la almohadilla retrodiscal de los traumatismos que produce el desplazamiento del complejo cndilo discal hacia atrs. Tambin limita la apertura rotacional y protege al
msculo pterigoideo lateral inferior de una excesiva distensin. Los ligamentos colaterales fijan el disco a la regin lateral y medial del cndilo mandibular y as el disco divide la articulacin en las cavidades supra e infra discai. Los ligamentos permiten la rotacin del cndilo mandibular bajo el disco, pero impiden o limitan el desplazamiento transversal, medial o Iateral del disco sobre el cndilo mandibular. El ligamento temporodiscal es uno de los responsables del desplazamiento medial del disco.
2.4 Membranas sinoviales
La superficie interna de la cpsula est tapizada por Ia membrana sinovial, la cual produce el lquido sinovial que se almacena en los fondos de saco de las cavidades supra e infradiscal. Las membranas sinoviales representan los medios de deslizamiento de la articulacin y estn formadas por dos capas: Ia sinovial ntima, que limita con los espacios de Ia articulacin, y la subsinovial unida al tejido conectivo fibroso de la cpsula. Estas membranas revisten por completo la cpsula articular del CATM adulto, tanto la cavidad superior, como inferior, pero estn ausentes en el tercio medio del disco en la articulacin adulto. La membrana sinovial contiene una poblacin heterognea de clulas. Entre ellas se destacan clulas con actividad fagoctica y clulas con capacidad de secrecin del cido hialurnico. Las clulas sinoviales aparecen dispuestas en una capa continua, aunque a menudo se hallan entremezcladas con fibras del conectivo capsular y con clulas adiposas. En ocasiones, forman vellosidades que se pueden proyectar hacia las cavidades de la articulacin. Algunas vellosidades son avasculares y otras contienen tejido conectivo y clulas adiposas. En general las vellosidades son escasas y aumentan en nmero en las patologas articulares. Con microscopia electrnica de transmisin se han identificado dos tipos de clulas sinoviales, tipo A y tipo B. Las clulas tipo A poseen un complejo de Golgi muy desarrollado y numerosas vesculas lisosomales, caracterstica de las clulas con actividad fagoctica. las tipo B poseen un complejo de Golgi ms pequeo, RER muy desarrollado y abundantes grnulos que producen una secrecin rica en
glicoprotenas y glicosaminoglicanos en re los que se destacan el cido hialurnico y la lubricina. Las clulas tipo A, menos abundantes( 20%) se originaran de los monocitos derivados de la mdula sea, en tanto que las clulas B (70%) se diferenciaran de las clulas mesenquimticas de los blastemas articulares( fig. 3). Recientemente se ha identificado la presencia de clulas dendrticas en el corion subsinovial, de funcin similar a las clulas de Merkel receptoras de sensaciones mecnicas (mecanoreceptores). La matiz extracelular (MEC) de la membrana sinovial contiene fibrillas de colgeno inmersas en un material amorfo electrodenso. L as clulas sinoviaIes estn ausentes en las zonas articulares funcionales. En Ia subsinovial (subntima) se pueden encontrar diversas variedades de tejido conectivo; de acuerdo a ello dicha capas e clasifican en: tipo areolar o laxa, tipo fibrosa y tipo adiposa (variedad ausente en condiciones normales).
2.5 Liquido sinovial
En las cavidadesarticularesexiste el lquido sinovial que tiene la funcin de lubricar y nutrir la articulacin. El lquido sinovial es producido como un ultrafiltrado del plasma sanguneo a partir de la rica red vascular de la membrana sinovial. Posee una coloracin amarillenta clara y contiene abundante cido hialurnico y mucinas, que le otorgan Ia viscosidad caracterstica. Tambin, presenta clulas libres descamadas y macrfagos. Normalmente, se deposita en los bordes y en el fondo de saco de la regin posterior. Durante los movimientos articulares, sin embargo, el lquido se desplaza de un sitio a otro (mecanismo conocido como lubricacin lmite). En reposo los sinoviocitos B elaboran pequeas gotitas de lquido sinovial para favorecer an ms Ia lubricacin articular (mecanismo llamado de lgrima). Desde el punto de vista funcional, el lquido sinovial tiene por finalidad, adems de lubricar las distintas regiones articulares, nutrir los condrocitos y por la capacidad fagoctica de los sinoviocitos A, degradar y eliminar las sustancias de desecho.
3. VASCULARIZACIN E INERVACIN
El CATM est bien vascularizado, porque posee un rico plexo vascular procedente de las arterias temporal superficial, timpnica anterior y farngea ascendente (ramas terminales de la cartida extema), que llegan hasta la cpsula articular Estas arterias se distribuyen en la periferia del disco, siendo la zona central avascular. Pequeos capilares se han encontrado en las vellosidades sinoviales subyacentes a la membrana sinovial. Dicha localizacin tiene importancia para la produccin del Iquido sinovial. El CATM est inervado por ramificaciones de Ios nervios auriculotemporal, masetero y temporal profundo, ramas del nervio trigmino, que pueden penetrar en la cpsula, disco y vellosidades sinoviales En la cpsula, las terminaciones nerviosas pueden ser del tipo de fibras nerviosas. Terminaciones nerviosas libres y encapsuladas (corpsculos de Rufini, Pacini y
10
Meissner). En el disco se observan slo terminaciones nerviosas libres (nocireceptores). En las vellosidades se han encontrado, tambin, terminaciones nerosas de aspecto corpuscular (mecanorecentores)
4. DESARROLLO DE LA ATM
4.1 Etapa inicial
Durante la 6a. semana embrionaria se observa diferenciacin del mesnquima vecino al cartlago de Meckel; en su extremo posterointerno se forman los mioblastos y miotbulos de la masa muscular pterigoidea y a partir de ellas los msculos pterigoideos interno y externo (1,2);en la parte externa se aprecia una condensacin o primordio articular. Durante la 7a. semana el primordio se diferencia en 2 blastemas: Temporal y condilar. Hacia el final de esta semana se condensa una lmina mesenquimal entre los blastemas. A la octava semana de gestacin, se identifican los blastemas condilar y glenoideo en el interior de una banda de ectomesnquima condensado que se
11
desarrolla adyacente al cartlago de Meckel y a la mandbula en formacin. Estos blastemas crecen a un ritmo diferente y se desplazan uno hacia el otro hasta enfrentarse a las 12 semanas.
El blastema condilar da lugar a la formacin del cartlago condilar, porcin inferior del disco y capsula articular. A partir del blastema glenoideo se forman la eminencia articular, regin posterosuperior del disco y porcin superior de la capsula Del tejido ectomesenquimal situado entre ambos blastemas se originan las cavidades supra intraarticulares. El cartlago primario de Meckel actuara como un componente de la actividad de ambos blastemas Existen evidencias que los huesecillos del odo medio, martillo y yunque, formados a partir del extremo posterior del cartlago de Meckel, funcionaran en el ser humano como una articulacin mvil hasta que se desarrolla el cndilo mandibular en relacin con la fosa mandibular del hueso temporal. e infradiscal, la membrana sinovial y los ligamentos
Entre la octava y la decimosexta semana aproximadamente, esta articulacin primaria seria funcional. Ms tarde, los cartlagos que forman el martillo y el yunque se osifican y quedan incorporados el odo medio. Los movimientos efectuados por esta articulacin primitiva y la contraccin muscular seran necesarios para asegurar una adecuada cavitacin articular La eminencia articular y la fosa mandibular adoptan su forma definitiva despus del nacimiento.
4.1.1 Desarrollo del cartlago condilar El cndilo, constituido por el cartlago condilar, es la estructura sobre la cual se ha puesto mayor nfasis por su participacin en el crecimiento mandibular. Durante mucho tiempo fue considerado un centro de crecimiento atribuyndosele la funcin
12
primordial de determinar la forma, tamao y ritmo de crecimiento de toda la mandbula Sin embargo, se ha demostrado que es un sitio de crecimiento porque es la mandbula a travs de los factores de crecimiento contenidos en los tejidos blandos que la rodean,la que controla y gua la forma del crecimiento condilar (teora de la matriz funcional de Moss) El cartlago condilar se encuentra unido a la parte posterior de la ramaascendente del cuerpo de la mandbula formado por cartlago hialinocubierto de una delgada
13
capa de tejido mesenquimtico fibroso Histolgicamente, en el cndilo de fetos humanos de diecisis semana hemos observado diversas zonas:
a) Zona superficial: est formada poruna cubierta mesenquimtica, cuya organizacin celular se asemeja a una membrana epiteloide (carece de lmina basal), sin embargo su estructura es tpicamente fibrosa con capilares en su interior. b) Zona Proliferativa: de mayor tamao que la anterior, estconstituida por clulas inmaduras que se encuentran incluidas en una densa red de fibras argirfilas y fibrillas colgenas .Estas clulas expresan la vimentina, marcador especfico del citoesqueleto de clulas mesenquimticas indiferenciadas. c) Zonas de condroblastos y condrocitos: estn constituidos porclulas cartilaginosas que se distribuyen al azar y se encuentran inmersasen una matriz extracelular (MEC) rica en proteoglicanos. d) Zona de erosin: se caracteriza por la presencia de condrocitoshipertrficos, MEC calcificada, clulas necrticas y condroclastos .En esta regin se observan tambin espculas seas delgadas en formacin ,con un patrn de distribucin no paralelo al eje del hueso en crecimiento ,como ocurre en la osificacin de los huesos largos .
Las trabculas seas de mayor tamao localizadas en la periferia del cuello del cndilo presentan una mayor radiopacidad. Las espculas centrales son ms pequeas, irregulares y estn constituidas principalmente por matriz osteoide escasamente mineralizada La envoltura externa del cndilo (pericondrio) se encuentra encontinuidad con la cubierta superficial mesenquimtica y con el periostioen diferenciacin.Las haces musculares del pterigoideo unidos a la superficie media delcndilo, estn formados por clulas musculares esquelticas quemuestran estriaciones
transversales tpicas.
14
1.1.2 Desarrollo del disco Articular
Alrededor de las doce semanas aparecen las cavidades infradiscal y supradiscal, que aparecen como una hendiduras en el ectomesnquima por encima y por debajo de Ia cabeza del cndilo. Los mecanismos que acontecen durante el
proceso de cavitacin an son desconocidos, sin embargo, en dicho proceso estaran involucrados mecanismos de apoptosis o de muerte celular programada, promovidos quizs, a partir de los movimientos del cndilo y de los tejidos conectivos adyacentes.
ANTES DEL NACIMIENTO En los fetos, el disco est formado por una banda delgada de tejido ectomesenquimtico con clulas semejantes a fibroblastos inmersas en una matriz rica en fibras argirfilas y escasas fibras colgenas. Su estructura bilaminar se hace evidente en eI rea retrodiscal, caracterizada por la presencia de abundantes y grandes vasos sanguneos y nervios.
15
Los extremos anterior y posterior del disco se extienden para constituir Ia cpsula, la cual est formada por un tejido conectivo menos fibroso, pero ms vascularizado e inervado. En el interior del disco, hemos identificado elementos nerviosos similares mecanoreceptores.
A medida que el desarrollo avanza, el cndilo, la fosa y el disco articular adquieren su contorno tpico, as, por ejemplo, el disco se observa delgado en la zona central y ms grueso en las regiones perifricas El tejido capsular que rodea a toda la articulacin, se extiende por delante hacia los haces musculares del pterigoideo y en la regin posterior se une al revestimiento mesenquimtico de la superficie del cndilo. En el interior de la cavidad articular el tejido conectivo de la superficie enva proyecciones que forman pliegues con pequeos capilares denominados vellosidades sinoviales. Los mioblastos que dan lugar a las fibras musculares del msculo pterigoideo externo, se forman a partir del mesnquima alrededor de la novena semana. Ms tarde las fibras musculares configuran dos haces: uno inferior que se fijar en el cndilo y otro superior que se unir al disco en formacin
16
ETAPA AVANZADA EN EL DESARROLLO DE LA ATM
Desde el punto de
vista anatmico, los componentes de la ATM quedan
establecidos aproximadamente en la decimocuarta semana de vida prenatal, aunque desde el punto de vista histofisiolgico son an estructuras inmaduras. A partir de este momento, los principales procesos que acontecen en el desarrollo de la ATM estn en relacin con la diferenciacin de los tejidos articulares, el aumento en las dimensiones de la articulacin y la adquisicin de su capacidad funcional. Con respecto a la maduracin neuromuscular buco facial, indispensable para alcanzar los reflejos de succin y deglucin que deben ejecutarse antes del nacimiento, se ha sugerido que comenzarn a partir de las catorce semanas de vida intrauterina, completndose alrededor de las veinte semanas Sato y colaboradores, mencionan que la maduracin condiIar y Ia diferenciacin de los msculos masticadores se produciran durante este perodo. El aumento de tamao del cndilo, se logra por mecanismos de crecimiento inrersticial y aposicional del cartlago condilar y por la formacin de trabculas seas mediante el proceso de osificacin endocondral, lo cual permite el crecimiento en longitud de la rama mandibular. La formacin de la fosa temporal comienza a las doce semanas con el desarrollo de gruesas trabculas seas por osificacin inrramembranosa. El tejido seo se contina formando ms all de las veintids semanas de vida prenatal paralelamente la fosa glenoidea desarrolla una pared media y otra lateral. La eminencia articular se diferencia entre las dieciocho y las veinte semanas, cuando Ia articulacin podra comenzar a ser funcional. El disco articular, aparece muy delgado en el rea central y engrosado en la periferia, donde se une a la cpsula articular, Ia cual a las veintisis semanas est completamente diferenciada. El disco en esta etapa, muestra una organizacin y distribucin especfica de las fibras colgenas y elsticas; dichas fibras se orientan en sentido antero posterior y tienden a aumentar con Ia edad.
17
En los ltimos meses del desarrollo prenatal, los cambios que ocurren estn principalmente relacionados con un aumento del tamao del cndilo y de Ia mandbula. El incremento en las dimensiones del maxilar inferior est ntimamente relacionado con Ia diferenciacin de los msculos masticadores. Estos msculos, junto a los factores de crecimiento presentes en los tejidos vecinos contribuiran al desarrollo del cndilo en la vida fetal. Las superficies articulares experimentan variaciones con la edad. Las los componentes seos incrementan paulatinamente en nmero, espesor y densidad. En el neonato, el disco est constituido por tejido Conectivo ricamente vascularizado. Sin embargo, en el desarrollo postnatal los vasos sanguneos
disminuyen considerablemente hasta convertir Ia regin central del disco adulto en una zona avascular y, persisten nicamente en los sitios de insercin.
18
4.2. DESARROLLO Y CRECIMIENTO POST- NATAL El crecimiento de la articulacin temporomandibular se contina hasta la segunda dcada de la vida postnatal. La morfologa del cndilo, la eminencia articular y de la fosa mandibular del temporal, adquieren su arquitectura tpica con la erupcin de los elementos dentarios. La fosa mandibular se profundiza y la eminencia articular se agranda a medida que se desarrollan los huesos laterales del crneo y aparecen los dientes primarios. Estas caractersticas anatmicas se acentan con la denticin permanente. La proliferacin de cartlago condilar y la formacin de tejido seo, es el que posibilita el crecimiento de la rama montante de Ia mandbula. Las superficies articulares y el disco, experimentan continuos cambios morfolgico para adaptarse a los nuevos requerimientos funcionales. La funcin articular es Ia que determina el crecimiento del cndilo y a su vez, su funcin depende del crecimiento y del desplazamiento mandibular. El aspecto histolgico del cndilo mandibular experimentan modificaciones con la edad. Es el tejido cartilaginoso el que, generalmente, proporciona la plasticidad de las superficies articulares.
19
Como puede verse en el cuadro 3, el cndilo del nio difiere del cndilo del adulto:
Entre los diecisiete y diecinueve aos la zona cartilaginosa se mineraliza y en sus capas profundas predominan los osteoclastos. Alrededor de los veintin aos, Ia amplitud de la capa proliferativa se reduce, lo que indica una disminucin en la tasa de crecimiento de Ia cabeza del cndilo y en consecuencia de la rama mandibular. Con la edad ocurre un cese definitivo de la actividad del cartlago condilar. Un hecho significativo sobre eI cartlago condilar comparado con otros cartlagos es que reacciona ms rpido y con un umbral ms bajo a los factores mecnicos extremos. Otro aspecto a destacar, es la diferencia que existe en la organizacin celular entre el cartlago condilar y el cartlago epifisiario de los huesos largos, tal como se muestra en el cuadro 4.
20
4.3. HISTOFISIOLOGA El normal funcionamiento de la ATM, permite que los movimientos mandibulares se realicen en las tres dimensiones del espacio de forma silenciosa, sin
interferencia y sin sensacin de molestia. En los movimientos masticatorios participan adems de los elementos dentarios, los msculos especficos y la ATM, regulados por guas seas, dentarias y sensoriales. Estas ltimas informan a travs de sus receptores del grado preciso de presin, para el correcto
funcionamiento de las estructuras comprometidas. Cualquier modificacin de la ATM o de la articulacin dentaria, puede provocar trastornos (o disfunciones) por su interdependencia funcional. Las caractersticas topogrficas de la articulacin Estn en estrecha relacin con la presencia o ausencia de los elementos dentarios y el tipo de dieta. Cuando se carece de piezas dentarias en boca, en las dos etapas extremas de la vida de un individuo, (lactantes y seniles) y Ia alimentacin predominante es de consistencia lquida o semislida, las superficies seas de la articulacin son poco profundas, en especial la fosa mandibular. En cambio, la existencia de dientes y una alimentacin mixta determinan anatmicamente el tpico aspecto de una diartrosis bicondlea.
21
Como se ha descrito anteriormente Ias estructuras articulares experimentan diversos tipos de cambios con Ia edad, como consecuencia de su adaptacin a diferentes condiciones funcionales. A partir de la etapa adulta, los tejidos estn sujetos al proceso natural de envejecimiento, lo que trae aparejado alteraciones tisulares y, por ende, disfunciones. Los cambios ms frecuentes encontrados en cada una de las estructuras de la ATM son los siguientes: Superficies articulares seas. A partir de los cincuenta y Cinco aos aproximadamente el cndilo, que est constituido por tejido seo, presentas signos de osteoporosis en diverso grado, siendo ms comn en la mujer (por ausencia de estrgenos) que en el hombre. Esta enfermedad que afecta a los huesos volvindolos frgiles por la movilizacin de Ca++, se manifiesta tambin, en de Ia mandbula y en el hueso temporal. A nivel de las superficies funcionales la cubierta fibrosa que acta como amortiguador fisiolgico junto con el disco, se vuelve de menor espesor.
Disco articular. Con la edad el disco presenta reas condroides especialmente en las zonas de mayor presin. Adems, puede observarse hialinizacin, acumulacin de agua y degeneracin de las fibras colgenas,
22
que constituyen un proceso irreversible, lo que conduce a Ia prdida progresiva de extensibilidad. En la regin retrodiscal, las paredes de los vasos aumentan de grosor. Membranas sinoviales. El nmero de vellosidades aumenta con la edad y particularmente en estados patolgicos (artrosis). Esto conlleva a una disminucin en la produccin de lquido sinovial y por ende, a una
reduccin en el nivel de lubricacin de las superficies articulares. Estas modificaciones son una de las causas de los ruidos o chasquidos articulares. Otras alteraciones que pueden presentar las membranas sinoviales estn en relacin con el aumento de clulas adiposas. Cpsula articular. En los individuos de edad avanzada, el tejido conectivo de la cpsula y de los ligamentos posee menor cantidad de capilares y nervios, volvindose fibroso, lo que limita los movimientos articulares. Msculos masticadores. Los msculos masticadores involucionan a partir de los sesenta y cinco aos, perdiendo considerablemente su eficacia funcional.
23
5.Sistema Nervioso
Los procesos de induccin, migracin y diferenciacin celular que se llevan a cabo durante la formacin del tejido nervioso generan un sistema altamente organizado capaz de proporcionar al nuevo ser una eficiente red de comunicacin con gran respuesta adaptativa y con la peculiaridad de responder autnomamente a estmulos fsicos y qumicos originados tanto en el medio interno como en interno. De esta manera, el sistema nervioso central (SNC) permite integrar y controlar las diferentes funciones del organismo.
Si se observa la evolucin de las especies, la centralizacin de la informacin es uno de los principios bsicos de la organizacin de los seres vivos, y es el SNC el encargado de asumir tales funciones. Un conocimiento bsico de la embriologa ayuda a comprender de mejor manera las intrincadas interrelaciones de los distintos componentes del SNC. 5.1 Desarrollo del Tubo Neural El sistema nervioso comienza su desarrollo embriolgico en la tercera semana (embrin de unos 1,5 mm.) en la lnea media de la regin dorsaldel embrin entre la membrana bucofarngea y el nodo primitivo.
Los procesos de formacin de la placa neural, pliegues neurales, y desarrollo del tubo neural se agrupan en el concepto de neurulacin.
24
Este perodo abarca desde el proceso de induccin notocordal hasta el cierre del neuroporo caudal. He aqu una breve descripcin de estos procesos: Al comenzar la tercera semana, la notocorda en desarrollo y el mesodermo
adyacente estimulan al ectodermo que est encima de ellos
Este complejo proceso de induccin notocordal hace que tejido ectodrmico (neuroectodermo) se engruese, formndose as la placa neural
25
. Actualmente, se han identificado varios tipos de molculas que actan como seales en los procesosde induccin y diferenciacin del SNC. As por ejemplo la interaccin entre BMP (bone morphogenetic protein), cordina y cido retonoico determinan la induccin y diferenciacin selectiva de ectoderma que forma piel de aquel que forma tubo neural ceflico y tubo neural mas caudal. La induccin neural trae como consecuencia una sobreproduccin inicial de clulas nerviosas. Se ha demostrado que a tal perodo prosigue otro de muerte celular programada o apoptosis, lo que determina la cantidad total de neuronas que el individuo tendr durante su vida. Tal nmero alcanza la impresionante cifra de alrededor de un
26
billn
de
neuronas.
Una vez completado el proceso inductivo, la placa neural se alarga desde su sitio de origen craneal al nodo primitivo hasta la membrana bucofarngea. Alrededor del da 18 del desarrollo los bordes laterales de la placa neural se elevan y forman los pliegues neurales.
La porcin media entre los pliegues neurales forma el Surco neural Hacia el final de la tercera semana los pliegues neurales se elevan an ms, se acercan y se fusionan irregularmente en la lnea media formando el tubo neural .
27
La fusin empieza en la regin cervical y sigue hacia ceflico y caudal. Mientras ocurre la fusin, los bordes libres del ectodermo superficial se separan del tubo neural. Posteriormente, ambos bordes se unen y forman una capa continua en la superficie que dar origen al epitelio epidrmico.Debido a que la fusin de los pliegues neurales no ocurre simultneamente a lo largo de ellos, la luz del tubo neural comunica con la cavidad amnitica en sus extremos ceflico y caudal a travs de los neuroporos craneal (anterior) y caudal (posterior). El cierre del neuroporo craneal se realizainicial el da 25 (perodo 18-20 somitos). El neuroporo caudal se cierra elda 27 (perodo de 25 somitos). El cierre de ambos neuroporos coincide con el establecimiento de la circulacin sangunea hacia el tubo neural.
28
Mientras los pliegues neurales se acercan a la lnea media para fusionarse, un grupo de clulas neuroectodrmicas ubicadas en la cresta de cada pliegue (cresta neural) pierden su afinidad epitelial con las clulas de la vecindad. La migracin activa de las clulas de la cresta neural desde las crestas hacia el mesodermo adyacente transforma el neuroectodermo en una masa aplanada e irregular que rodea al tubo neural. Este grupo celular dar origen a un conjunto heterogneo de tejidos de gran importancia: ganglios de la raz posterior, ganglios autnomos, ganglios de los pares craneales V, VII, IX, X, clulas de Schwann, las leptomeninges (aracnoides y piamadre), melanocitos, mdula suprarrenal, odontoblastos. En consecuencia, el tubo neural ser el que se convertir por diferenciacin en encfalo y mdula espinal, mientras que las crestas neurales formarn la mayor parte del sistema nervioso perifrico (SNP) y parte del autnomo (SNA).
29
Luego del cierre completo del tubo neural, comienza el desarrollo de la regin caudal del tubo (segmentos sacros bajos y coccgeo) mediante procesos de canalizacin y diferenciacin regresiva. Entre el 28 y 32 da, un conjunto de clulas indiferenciadas al final del tubo neural desarrolla una serie de pequeas vacuolas que posteriormente toman contacto con el canal central. Es frecuente la presencia de innumerables canalculos que pudiesen ser causas de posteriores defectos. Este proceso de canalizacin prosigue hasta la sptima semana. En el periodo siguiente ocurre la regresin de muchas clulas de la masa caudal. Como remanentes de este proceso de diferenciacin regresiva quedan el ventrculo terminal y el filum terminale. El extremo ceflico del tubo neural se dilata y origina 3 vesculas enceflicas primarias: -Prosencfalo (cerebro anterior) -Mesencfalo (cerebro medio ) -Rombencfalo (cerebro posterior)
30
El tercio caudal del tubo se alarga y su dimetro se acorta para formar la mdula espinal. El neurocele se estrecha y pasa a formar el canal central (del epndimo) de la mdula espinal que se contina con la cavidad de las vesculas enceflicas. La cavidad del rombencfalo es el Cuarto ventrculo, la del diencfalo el Tercer ventrculo y la de los hemisferios cerebrales los Ventrculos laterales. Tercer y cuarto ventrculos se comunican por la luz del mesencfalo que se torna estrecha y origina el Acueducto cerebral (de Silvio). Los ventrculos laterales se comunican con el Tercer ventrculo por los agujeros interventriculares (de Monro) 5.2 Mdula Espinal Luego de ocurridos los sucesos de neurulacin, el tubo neural forma una estructura totalmente separada de la cavidad amnitica cuya pared est constituida por clulas cilndricas que la recorren formando un epitelio pseudoestratificado y que estn conectadas por complejos de unin (znula occludens, znula adherens y desmosomas hacia el neurocele.
Durante el perodo de surco neural incluso hasta despus de cerrarse el tubo neural, estas clulas proliferan para formar la Capa neuroepitelial (neuroepitelio). Esta capa da origen a todas las neuronas y neuroglias (astrocitos y oligodendrocitos) de la mdula espinal. Durante la quinta semana, las clulas neuroepiteliales proliferan y producen un aumento en longitud y dimetro del tubo neural. Adems, es posible observar cambios en la conformacin de los diferentes elementos intracelulares, como por ejemplo, modificaciones en la morfologa del ncleo o la presencia de un mayor nmero de ribosomas asociados al retculo endoplsmico consecuencia del considerable aumento en la actividad
neurosecretora. Estas clulas denominadas Neuroblastos (clulas nerviosas primitivas) migran a la periferia y se organizan en una nueva estructura: la Capa del manto, la que posteriormente constituir la sustancia gris de la mdula espinal.
31
Las prolongaciones axonales de las neuronas de la capa del manto migran a la periferia y forman los fascculos nerviosos de la Capa marginal. Al mielinizarse estas fibras nerviosas, la capa toma un aspecto blanquecino y constituye la sustancia blanca de la mdula espinal
Gran parte de los glioblastos (clulas de sostn primitivas) deriva del neuroepitelio una vez que este ya ha dado origen a los neuroblasto.
Los glioblastos emigran desde la capa neuroepitelial hacia las capas marginal y del manto para all diferenciarse en Astrocitos tipo I (aquellos que envan prolongaciones a la piamadre y a los vasos sanguneos enceflicos para formar la barrera hematoenceflica) y tipo II (que toman contacto con los nodos de Ranvier de los nervios mielnicos del SNC y suelen encapsular las sinapsis qumicas). Los astrocitos tienen importantes funciones en el SNC: 1) Forman la barrera hematoenceflica que protege al SNC de cambios bruscos en la concentracin de iones del lquido extracelular y de otras molculas que pudiesen interferir en la funcin neural. 2) Eliminan K+, glutamato y cido, gama-aminobutrico del espacio
32
extracelular. 3) Son importantes almacenes de glucgeno; realizan glucogenlisis al ser inducidos por norepinefrina o pptido intestinal vasoactivo (VIP). La clula de oligodendroglia son glias del SNC que forman las vainas de mielina que rodean a los axones de los tractos del SNC. Los oligodendrocitos satlites son aquellos que rodean los cuerpos celulares y regulan el ambiente bioqumico que rodea la neurona.
Alrededor del cuarto mes aparecen las clulas de microglia. Derivan del mesnquima circundante y se caracterizan por ser pequeas y muy fagocticas. Llegan a la sustancia blanca y gris del SNC luego de la aparicin de los vasos sanguneos. Cuando las clulas neuroepiteliales dejan de producir neuroblastos y glioblastos, se diferencian las clulas ependimarias que revisten el canal central de la mdula espinal.
5.3 Desarrollo de las placas basales, alares, del techo y del piso
La multiplicacin de los neuroblastos de la capa del manto a cada lado del tubo neural origina unos engrosamientos en la regin ventral y dorsal:
1)Las placas basales engrosamiento ventral) incluyen los somas de las motoneuronas que posteriormente constituirn las astas anteriores de la mdula espinal. 2)Las placas alares (engrosamientos dorsales) corresponden a regiones sensitivas que se diferenciarn en las astas posteriores de la mdula espinal.
El crecimiento de las placas alares origina el tabique medio posterior. Al sobresalir ventralmente las placas basales se forma el tabique medio anterior, mientras tanto se desarrolla la fisura mediana anterior en la superficie anterior de la mdula espinal. El surco limitante delimita ambas placas, y de esta manera tambin separa las regiones motoras de las sensitivas. Las regiones dorsal (placa del
33
techo) y ventral (placa del piso) en la lnea media del tubo neural no poseen neuroblastos y constituyen vas para fibras nerviosas que cruzan la mdula espinal de un lado al otro.
Entre las astas ventral y dorsal de los segmentos torcicos hasta el segundo o tercero lumbar de la mdula espinal se acumulan neuronas que formarn el asta lateral o intermedia, que contiene neuronas del Sistema nervioso autnomo. 5.4 Las meninges espinales y las modificaciones durante la formacin de la mdula espinal El tejido mesenquimtico (esclerotoma) que rodea el tubo neural se condensa para formar la meninge primitiva, que originar la duramadre. A esta meninge primitiva se le agragan clulas provenientes de las crestas neurales para formar la capa interna denomianda leptomeninges (aracnoides y piamadre). Al unirse los espacios llenos de lquidos que existen entre las leptomeninges, se forma el espacio subaracnodeo. El origen de la aracnoides y piamadre a partir de una capa nica explica la existencia de las trabculas aracnoideas que existen entre ellas. Como resultado del desarrollo del aparato locomotor durante el cuarto mes, adems de la adicin de neuronas motoras y sensitivas, la mdula espinal se ensancha en las regiones cervical y lumbar formando los engrosamientos cervical y lumbar.
Al tercer mes, la mdula espinal se extiende a lo largo del canal vertebral del embrin y los nervios espinales atraviesan los agujeros intervertebrales a nivel de su origen. Poco despus, la columna vertebral y la duramadre se alargan ms rpido que el tubo neural ocasionando que el extremo terminal de la mdula se desplace a niveles ms altas.
34
A los seis meses de vida intrauterina alcanza la primera vrtebra sacra, y ya en el neonato su extremo est a nivel de L3. Debido a este crecimiento desproporcionado, los nervios raqudeos tienen una direccin oblicua desde su segmento de origen en la mdula espinal hasta el nivel correspondiente de la columna a nivel coccgeo.
En el adulto, la mdula espinal termina a nivel L2 (esta es una medida promedio, ya que el extremo medular puede estar tan alto como T12 o tan bajo como borde superior de L3). Debajo, una prolongacin filiforme de la piamadre forma el filum terminale que se adosa al periostio de la primera vrtebra coccgea y seala la lnea de regresin de la mdula espinal embrionaria. Las fibras nerviosas bajo el extremo inferior de la mdula espinal forman la Cauda equina, cuya denominacin se debe a su semejanza a la cola de caballo. Cuando se extrae LCR por una puncin lumbar, la aguja se introduce en un nivel lumbar bajo respetando as el extremo terminal de la mdula espinal
35
5.5 Encfalo Las estructuras enceflicas aparecen luego de ocurridos cuatro procesos bsicos: (a) proliferacin neuronal (b) migracin (c) perodo de organizacin, el cual se establece la diferenciacin celular. Este se desarrolla hasta el nacimiento una vez una vez establecido el patrn de funcionamiento de las diferentes regiones enceflicas, y (d) mielinizacin.
Durante la cuarta semana, despus del cierre de los neuroporos, el extremo ceflico del tubo neural craneal al cuarto par de somitos se dilata
considerablemente y aparecen las tres vesculas enceflicas primarias a partir de las cuales se origina el encfalo: Prosencfalo (cerebro anterior), Mesencfalo (cerebro medio) y Rombencfalo (cerebro posterior). Simultneamente se estn formando dos plegamientos: el pliegue cervical, en la unin del rombencfalo y mdula espinal y el pliegue ceflico en el mesencfalo. El mesencfalo dista del rombencfalo por un surco: el istmo del rombencfalo (de His)
Durante la quinta semana el prosencfalo y rombencfalo se dividen en dos vesculas secundarias: El prosencfalo origina (1) el Telencfalo (cerebro terminal) que consta de una parte media y dos evaginaciones laterales (hemisferios cerebrales primitivos) (2) el Diencfalo, que presenta la evaginacin de las vesculas pticas. El rombencfalo formar finalmente (1) el Metencfalo, que constituir la protuberancia y el cerebelo (2) el Mielencfalo, el futuro bulbo raqudeo. El lmite metencfalo-mielencfalo queda definido por el pliegue protuberancial.Este pliegue se origina debido al crecimiento desigual del rombencfalo dando lugar a un adelgazamiento de su techo.
Al principio, el encfalo tiene su estructura bsica muy similar a la mdula espinal, sin embargo, debido a la aparicin de los pliegues y surcos enceflicos se producen variaciones considerables en la disposicin de los diferentes elementos. En general, las placas alares y basales del rombencfalo y mesencfalo se encuentran bien definidas, en cambio, en el prosencfalo las placas alares estn acentuadas y las basales en plena regresin.
36
5.6 ROMBENCEFALO Est formado por: (1) Mielencfalo (2) Metencfalo MIELENCEFALO Es la vescula enceflica ms caudal y se diferencia en el bulbo raqudeo (mdula oblonga) Sus paredes laterales sufren cierta eversin tal como se abren las conchas de una almeja, sin embargo, su estructura general es bastante parecida a la mdula espinal.
Los neuroblastos de las placas alares migran a la capa marginal en direccin ventrolateral para formar los ncleos olivares Ventralmente, las fibras
corticospinales que descienden desde la corteza cerebral (giro precentral) forman las denominadas pirmides. El pliegue protuberancial hace que las paredes
37
bulbares laterales se desplacen lateralmente y que la placa del techo se extienda y adelgace considerablemente. Como consecuencia, la cavidad del mielencfalo (futuro IV ventrculo) toma forma romboide y los ncleos motores pasan a ubicarse medialmente a los ncleos sensitivos.
Las placas alares y basales estn bien definidas. La placa basal contiene 3 grupos de ncleos motores: (1) Eferente somtico o medial (2) Eferente visceral especial o intermedio (3) Eferente visceral general o lateral. Estos tres grupos originan los ncleos motores de los nervios craneales IX, X, XI y XII que se ubican en el piso del cuarto ventrculo medial al surco limitante. La placa alar contiene tres grupos nucleares sensitivos: (1) Aferente somtico o lateral (2) Aferente visceral especial o intermedio (3) Aferente visceral general o medial. Estos grupos neuronales forman los ncleos sensitivos de los nervios craneales V, VII, VIII, IX y X y los ncleos gracilis y cuneatus
38
5.7 METENCEFALO
Incluye la regin ubicada entre el pliegue protuberancial y el istmo del rombencfalo. La porcin metenceflica ventral ms una contribucin celular de la regin alar del mielencfalo originan el Puente (protuberancia), mientras la regin posterior conforma el cerebelo. El puente forma una importante va nerviosa entre la mdula espinal y las cortezas cerebral y cerebelosas. Por otra parte, el cerebelo es un centro de coordinacin de postura y movimientos. La cavidad del metencfalo forma la parte superior del futuro IV ventrculo.
La formacin del pliegue protuberancial produce el distanciamiento de las paredes laterales del puente y la extensin de la sustancia gris del piso del IV ventrculo. Los neuroblastos de las placas basales constituyen tres columnas de ncleos motores: (1) Eferente somtico medial (2) Eferente visceral especial (3) Eferente visceral general. Ellos originan los ncleos motores de los pares V, VI y VII. La capa marginal de las placas basales se expande y sirve de puente a fibras que conectan la mdula espinal con las cortezas cerebral y cerebelosas; esto explica el nombre de "puente".
Las placas alares poseen 2 grupos sensitivos (1) Aferente somtico lateral (2) Aferente visceral general.
39
Ellos constituyen el ncleo sensitivo principal del n.trigmino, el ncleo espinal del V par y los ncleos vestibulares del VIII par. Los ncleos pontinos se originan en las placas alares del metencfalo.
5.7.1 Cerebelo: Cada placa alar se curva en su regin dorsolateral en direccin medial para formar los labios rmbicos.Estos labios aumentan de tamao, se proyectan
caudalmente sobre la placa del techo del IV ventrculo y se fusionan en la lnea media. En la zona inferior del metencfalo estn muy separados. La compresin cefalocaudal de los labios producto de la exageracin del pliegue protuberancial forma la placa cerebelosa que se superpone al puente y al bulbo raqudeo. En el embrin de 12 semanas se observa una parte media (vermis) y dos laterales (hemisferios) Inicialmente, la placa cerebelosa consta de las capas neuroepitelial, del manto y marginal, pero luego algunas clulas neuroepiteliales emigran a la superficie cerebelosa a formar la capa granulosa externa que consta de una zona proliferativa superficial. Al sexto mes, la capa granulosa externa ya ha producido clulas granulosas, clulas en cesto y clulas estrelladas que contactan con clulas de Purkinje an indiferenciadas. La corteza cerebelosa alcanza sus dimensiones definitivas despus del nacimiento. (emboliforme, globoso y Los ncleos dentados y dentados accesorios se presencian antes del nacimiento.
fastigio)
Posteriormente, los axones que salen de estos ncleos cruzan el mesencfalo para llegar al prosencfalo y constituyen el pednculo cerebeloso superior. El crecimiento axonal de las fibras corticopontinas y pontocerebelosas que conectan las cortezas cerebral y cerebelosa conlleva la formacin del pednculo cerebeloso medio. Axones sensitivos provenientes de la mdula espinal, ncleos olivares y vestibulares forman el pednculo cerebeloso inferior.
40
5.8 MESENCEFALO
El mesencfalo constituye la vescula enceflica inmediatamente ceflica al rombencfalo que sufre menos modificaciones durante el desarrollo del SNC. La cavidad de la vescula mesenceflica se reduce considerablemente para formar un conducto que unir los futuros III y IV ventrculos: el acueducto cerebral (de Silvio). A cada lado, las placas basales y alares estn separadas por el surco limitante. Cada placa basal tiene 2 grupos de motoneuronas (1) Eferente somtico o medial: origina los nervios craneales III y IV (2) Eferente visceral general: forma el ncleo de Edinger-Westphal. La capa marginal de las placas basales se expande y origina el pie de los pednculos cerebrales por donde descienden fibras desde la corteza cerebral a centros motores inferiores del puente y mdula espinal (tractos corticopontinos, corticobulbares y corticoespinales). Las placas alares y del techo forman el
41
tectum. Neuroblastos de las placas alares migran a la capa marginal del tectum y forman agregados estratificados de neuronas sensitivas separadas por un surco transverso: los colculos superiores (anteriores) y los colculos inferiores (posteriores). Estos ltimos son centros de relevo para reflejos auditivos, mientras que los colculos anteriores forman centros de correlacin y de reflejos para estmulos visuales.
Una banda de sustancia gris adyacente al pednculo cerebral forma la sustancia negra (locus niger). Se tienen muchas dudas si este elemento se origina a partir de clulas de las placas basales o de clulas de las placas alares que migran ventralmente. 5.8.1 PROSENCEFALO Antes del cierre del neuroporo craneal, aparecen dos divertculos laterales a cada lado del diencfalo: las vesculas pticas (primordios de la retina y nervios pticos). Posteriormente, aparecen las vesculas telenceflicas que formarn los hemisferios cerebrales y sus cavidades: los ventrculos laterales. Las placas del piso y del techo son delgadas; por otra parte, las paredes laterales son gruesas como en la mdula espinal. 5.8. 3 DIENCEFALO Se desarrolla a partir de la porcin media del prosencfalo y consta de placas del techo y alares pero carece de placas basales y del piso. El mesnquima vascularizado de los ependimocitos de la placa del techo origina el plexo coroideo del tercer ventrculo. La porcin caudal de la placa del techo forma un divertculo ubicado anteriormente al mesencfalo que hacia la sptima semana ya forma un rgano macizo con forma de cono: el cuerpo pineal (epfisis). El calcio acumulado en la epfisis durante la adultez la hace un importante punto de referencia en imagenologa. Tiene importantes funciones sexuales (secrecin de melatonina que estimula la secrecin de gonadotropinas) y es posible que forme un nexus entre los ciclos de luz solar y endocrinos (ritmos circadianos) y de la
42
conducta. En las paredes laterales del Tercer ventrculo (placas alares del diencfalo) aparecen tres prominencias que posteriormente formarn el hipotlamo tlamo y epitlamo . El surco hipotalmico divide las placas alares en una porcin ventral (hipotlamo) y una dorsal (tlamo). Este surco no es la continuacin del surco limitante, como se creia antiguamente, y no divide porciones sensitivas y motoras. La notable proliferacin ocurrida en el tlamo hace que ste protruya hacia el III ventrculo de modo que las regiones talmicas derecha e izquierda se fusionan en la lnea media formando la adhesin intertalmica (presente en un 70 a 80% de los cerebros).
El hipotlamo (porcin inferior de la placa alar) se diferencia en varios grupos nucleares que constituyen centros reguladores de variadas funciones del organismo (temperatura corporal, emociones, hambre, saciedad, sueo, etc.). Uno de estos ncleos, los cuerpos mamilares, sobresalen en la superficie ventral del hipotlamo a cada lado de la lnea media.
El techo y la parte dorsal de la pared lateral del diencfalo formarn el epitlamo. Desarrollo de la Hipfisis (glndula pituitaria): La hipfisis se origina totalmente del ectodermo (cuarta semana). Se desarrolla a partir de dos porciones: (1) Una evaginacin dienceflica hacia caudal (Infundbulo). (2) Una evaginacin ectodrmica del estomodeo (cavidad bucal primitiva) anterior a la membrana bucofarngea (Bolsa de Rathke). Este doble origen explica la diferencia de tejidos hipofisiarios. En la tercera semana, la bolsa de Rathke crece dorsalmente hacia el infundbulo. Al final del segundo mes, pierde contacto con la cavidad bucal y se contacta ntimamente con el infundbulo. Posteriormente, la multiplicacin de las clulas de la pared anterior de la bolsa de Rathke originan el lbulo anterior de la hipfisis (Adenohipfisis). La adenohipfisis consta de tres partes: (1) Una prolongacin de este lbulo que crece rodeando el tallo del infundbulo: la pars tuberalis. (2) la pars distalis que constituye el lbulo anterior propiamente tal.
43
(3) la pared posterior de la bolsa de Rathke no prolifera y forma la pars intermedia, de poca importancia en el humano.
La evaginacin dienceflica origina la eminencia media, el tallo infundibular y la pars nerviosa que en conjunto se les denomina Neurohipfisis (lbulo posterior). El extremo distal del neuroepitelio del infundbulo experimenta una proliferacin que origina los pituicitos, las clulas de sostn de la neurohipfisis.
La proliferacin de la pared anterior de la bolsa de Rathke reduce su luz hasta formar una pequea hendidura (ella es un posible sitio de quistes, por ello, es importante conocerla). 5.9 TELENCEFALO
Es la vescula enceflica ms rostral. Consta en 2 evaginaciones laterales (hemisferios cerebrales) y una porcin media (lmina terminal). Sus cavidades (ventrculos laterales) comunican con el III ventrculo a travs de los agujeros interventriculares. Los Hemisferios Cerebrales: Entre la 5 y12 semana, las evaginaciones bilaterales de la pared lateral del telencfalo originan los hemisferios cerebrales. La expansin anterior forma los lbulos frontales mientras la superolateral origina los lbulos parietales; finalmente, la expansin posteroinferior forma los lbulos temporales y occipitales. El proceso contina con un aplanamiento medial de los hemisferios cerebrales.
El mesnquima de la fisura longitudinal del cerebro origina la hoz del cerebro (falx cerebri). La pared medial de los hemisferios, donde se unen al techo dienceflico, es delgada y slo consta de una capa de clulas ependimarias cubierta de mesnquima vascularizado: el plexo coroideo del tercer ventrculo.
Durante la 6 semana, la parte basal de los hemisferios aumenta de tamao y sobresale hacia el ventrculo lateral dando origen al Cuerpo estriado. Esta regin
44
de la pared hemisfrica se expande en direccin posterior y se divide en 2 partes: (1) ncleo caudado, dorsomedialmente (2) ncleo lenticular, ventrolateralmente. La fusin de la pared medial del hemisferio y la pared lateral del diencfalo permite el contacto entre el ncleo caudado y tlamo. Tractos ascendentes y descendentes de la corteza cerebral pasan entre tlamo y ncleo caudado medialmente y ncleo lentiforme lateralmente formando la cpsula interna. La pared del prosencfalo se engrosa formando una estructura longitudinal que protruye al ventrculo lateral: el hipocampo.
La zona suprayacente al ncleo lentiforme crece lentamente y queda ocultada entre los lbulos temporal y occipital (lbulo de la nsula). Al final de la vida fetal, la superficie hemisfrica crece tan rpido que se forman giros (circunvoluciones) separados por surcos y cisuras. Estos surcos y giros permiten un aumento considerable de la superficie cerebral y, por ende, un aumento de la superficie cortical sin sobrepasar el volumen del crneo. 5. 10 La Corteza Cerebral:
La corteza cerebral se desarrolla a partir del palio, que consta de tres regiones: (1) Paleopalio, (2)Arquipalio, (3) Neopalio
Estas originan la paleocorteza, la arquicorteza y la neocorteza respectivamente. En cualquier region de la corteza, las paredes de los hemisferios cerebrales presentan tres zonas: (1) ventricular (2) intermedia (3) marginal, ms una que se agrega ulteriormente (zona subventricular).
Las neuronas que migran desde la zona intermedia (regin subependimaria) hacia la zona marginal originarn la corteza cerebral
45
. De esta manera, la sustancia gris queda ubicada superficialmente y los axones o fibras nerviosas quedan ubicadas en la profundidad del cerebro. La primera masa de neuroblastos que emigra en el neopalio se dirige a una zona inmediatamente debajo de la piamadre para diferenciarse en neuronas maduras. Las siguientes oleadas de neuroblastos van ubicndose entre la piamadre y la capa anteriormente formada. En conclusin, los primeros neuroblastos formados quedan en la porcin profunda de la corteza mientras que los formados posteriormente originan las capas superficiales de la corteza. La diferenciacin neuronal en las diferentes capas da un aspecto estratificado a la corteza cerebral y origina zonas con una composicin celular especfica. Por ejemplo, las clulas piramidales abundan en la corteza motora y las clulas granulosas se encuentran en gran cantidad en las regiones sensitivas.
46
5. 11 Comisuras Las comisuras cerebrales son un grupo de axones que atraviesan la lnea media a diferentes niveles y conectan los hemisferios cerebrales derecho e izquierdo La lmina terminal (extremo ceflico del tubo neural) se extiende desde la placa del techo del diencfalo hasta el quiasma ptico.
La estructura comisural ms importante que abarca gran parte de las fibras del sistema comisural de la corteza cerebral es el cuerpo calloso. Se desarrolla durante la 10 semana como un pequeo fascculo en la lmina terminal y comunica regiones no olfatorias de ambos hemisferios. La expansin del neopalio hace que el cuerpo calloso crezca hacia anterior y luego posteriormente rebosando la lmina terminal y formando un arco sobre el techo dienceflico. Todo este proceso induce un estiramiento de la lmina terminal entre el cuerpo calloso y el frnix que origina el septum pellucidum. La comisura anterior constituye la primera comisura en formarse y conecta la corteza temporal y el bulbo olfatorio de un lado y otro. Se ubica superiormente a la lmina terminal. El frnix nace en el hipocampo, converge en la lmina terminal y prosigue hacia posterior hasta llegar a los cuerpos mamilares y al hipotlamo. La comisura posterior, la comisura habenular y el quiasma ptico tambien son estructuras que permiten la pasada de axones hacia el lado opuesto del cerebro.
47
5.12 Proceso de Mielinizacin: Comienza durante el periodo fetal tardo y generalmente contina durante los dos primeros aos de vida postparto. La mielinizacin de nervios perifricos la realizan las clulas de Schwann que migran a la periferia y se disponen alrededor de los axones formando la Vaina de Schwann (antiguamente denominada neurilema). Durante el 4 mes, muchas fibras nerviosas toman aspecto blanquecino por el depsito de mielina que se forma por el repetido enrollamiento de la membrana de la clula de Schwann alrededor del axn.
La mielinizacin de las fibras de la mdula espinal comienza en el cuarto mes de vida prenatal desde la regin cervical hacia caudal, aunque algunas fibras nerviosas que vienen desde centros cerebrales superiores hacia la mdula no se mielinizan sino que hasta periodos de vida postnatal. La vaina de mielina que rodea las fibras nerviosas de la mdula espinal tiene su origen en las clulas de oligodendroglia. Las fibras de las races posteriores se mielinizan despus que lo hacen las races anteriores, por tanto son las fibras funcionalmente motoras las que realizan el proceso de mielinizacin en primer lugar.
En el cerebro, el proceso mielinizante comienza en la sexta semana de vida fetal en las fibras del cuerpo estriado. Las fibras sensitivas que suben al encfalo desde la mdula espinal son las segundas en mielinizarse. La mielinizacin del encfalo es tan lenta que al nacimiento slo una pequea porcin ha completado el proceso. Aquello se refleja en una pobre capacidad motora del recin nacido, cuyas principales acciones involucran en su mayora reflejos. En el perodo postnatal, la mielinizacin se vuelve sistemtica y se realiza en diferentes regiones en tiempos especficos. Por ejemplo, es sabido que las fibras del tracto piramidal se mielinizan en la sexta semana de vida postnatal. Investigaciones recientes muestran que algunas fibras enceflicas no se mielinizan sino hasta la pubertad. Se cree que los tractos del sistema nervioso se mielinizan al adquirir su capacidad funcional.
48
5.13 Plexos coroideos y Lquido Cefalorraqudeo (LCR): El epndimo que se encuentra en el techo del IV ventrculo est recubierto externamente por la piamadre. En conjunto, estas estructuras forman la tela coroidea. La proliferacin de las clulas piales provoca una invaginacin hacia el IV ventrculo de la tela coroidea (plexo coroideo del IV ventrculo). Similar fenmeno ocurre en el III ventrculo y en las fisuras corodeas de los ventrculos laterales. La funcin de los plexos coroideos es la secrecin del lquido cefalorraqudeo (LCR), hacia el sistema ventricular. La absorcin del LCR se realiza hacia el sistema venoso a travs de las vellosidades aracnoideas. Estas vellosidades son protrusiones de la aracnoides hacia los senos venosos de la duramadre, y consisten en una delgada capa de clulas que deriva del endotelio de los senos venosos y del epitelio aracnoideo.
49
50
6. DESARROOLLO DEL SISTEMA NERVIOSO PERIFERICO
El sistema nervioso perifrico (SNP) consta de nervios craneales, raqudeos y viscerales, as como los ganglios craneales, raqudeos y autnomos. El SNP se desarrolla a partir de diversas fuentes, fundamentalmente de la cresta neural. Todas las clulas, fundamentalmente de la cresta neural. Todas las clulas sensitivas (somticas y viscerales) del SNP proceden de las clulas de la cresta neural. Los cuerpos celulares de estas clulas sensitivas se localizan fuera del SNC. Con excepcin de las clulas del ganglio espiral de la cclea y el ganglio vestibular del PC VIII (nervio vestibulococlear), todas las clulas sensitivas perifricas son inicialmente bipolares, pero ambas prolongaciones se unen pronto para formar una sola y un tipo de neurona unipolar (fig. 1).
Fig. 1: Diagrama de fases sucesivas de la diferenciacin de una clula de la cresta neural en una neurona aferente unipolar de un ganglio raqudeo.
Esta prolongacin tiene unas ramificaciones perifrica y central. La ramificaron perifrica finaliza en una terminacin sensorial, mientras que el central penetra en la medula espinal o el encfalo (Fig. 2). Las clulas sensitivas del ganglio del PC VIII conservan la bipolaridad. El cuerpo celular de cada neurona aferente se recubre de una capsula de clulas de Schwann modificadas, las
51
clulas satlite (Fig. 2), que proviene de clulas de la cresta neural. Esta capsula es continua con la vaina neurolemal de las clulas de Schwann que rodea los axones de las neuronas aferentes. En la parte externa de las clulas satlite existe una capa de tejido conjuntivo que se contina con una vaina endoneural de las fibras nerviosas. Este tejido conjuntivo y las vainas endoneurales proceden del mesnquima.
Fig. 2: Diagrama que indica los derivados de la cresta neural. Las clulas de la cresta neural tambin se diferencian en las clulas de los ganglios aferentes de los pares craneales y muchas otras estructuras. Asimismo se ilustra la formacin de un nervio raqudeo.
52
Las clulas de la cresta neural en el encfalo en desarrollo migran para formar ganglios sensoriales solamente en relacin con los nervio trigmino (PC V), facial (PC VII), vesticulococlear (PC VIII), glosofarngeo (PC IX) y vago (PC X). Las clulas de dicha cresta tambin se diferencian en neuronas multipolares de los ganglios autnomos (Fig. 3), incluyendo ganglios de los troncos simpticos que se encuentran a los lados de los cuerpos vertebrales; ganglios colaterales o prevertebrales en plexos del trax y abdomen (como los plexos cardiaco, celiaco y mesentrico), y ganglios parasimpticos o terminales en o cerca de las vsceras (p.ej. plexos submucoso o de Meissner). Las clulas de los paraganglios o clulas cromafines tambin derivan de la cresta neural. El termino paraganglio abarca varios grupos muy diseminados de clulas similares en varios respectos a las medulares de las glndulas suprarrenales. Los grupos celulares se hayan principalmente en posicin retroperitoneal, con frecuencia en asociacin con los ganglios simpticos. Los cuerpos carotideos y articos tambin poseen pequeas islas de clulas cromafines asociadas a ellos. Estos grupos diseminados de clulas cromafines constituyen el sistema cromafin. As mismo, las clulas de la cresta dan lugar a los melanoblastos (precursores de los melanocitos) y a las clulas de la medula de la glndula suprarrenal.
6.1 Nervios raqudeos
Las fibras nerviosas motoras que surgen de la medula espinal comienzan a aparecer a finales de la cuarta semana. Las fibras nerviosas salen de clulas de las placas basales de la medula espinal en desarrollo y emergen como una serie continua de raicillas a lo largo de su superficie ventrolateral. Las fibras destinadas a un grupo muscular determinado en desarrollo se organizan en un haz, formando una raz nerviosa ventral. Las fibras nerviosas de la raz nerviosa dorsal estn compuestas por axones procedentes de las clulas de la cresta neural que migran
53
hacia la cara dorsolateral de la medula espinal, donde se diferencian en clulas de los ganglios raqudeos (Fig. 3).
Fig. 3: Microfotogrfica de un corte transversal de la medula espinal en desarrollo en un embrin humano en el estadio 20 de Carnegie de alrededor de 50 das (x60)
Las prolongaciones centrales de las neuronas de los ganglios raqudeos forman un nico haz que crece hacia la medula espinal contigua al vrtice del asta dorsal de la sustancia gris (fig. 4).
54
Fig. 4: Diagramas que ilustran el desarrollo de la medula espinal, B y C, Cortes similares ala sexta y novena semana respectivamente.
Las prolongaciones dstales de las clulas de los ganglios raqudeos crecen haca la raz nerviosa ventral y finalmente se unen a ella para formar un nervio raqudeo. Inmediatamente antes de su formacin, un nervio raqudeo mixto se divide en ramas primarias dorsal y ventral (L. Rami, ramas). La rama primaria dorsal, la divisin menor, inerva la musculatura axial dorsal (Fig. 5), vrtebras, articulaciones intervertebrales posteriores y parte de la piel de la espalda. La rama primaria ventral, divisin mayor de cada nervio raqudeo, participa en la innervacin de las extremidades y partes ventrolaterales de la pared corporal. Los plexos nerviosos mayores (cervical, humeral y lumbosacro) estn formados por ramas primarias ventrales. A medida que se desarrolla cada yema de la extremidad, los nervios de los segmentos de la mdula espinal opuestos a estas se alargan y crecen hacia el miembro. Las fibras nerviosas se distribuyen hasta sus msculos, que se diferencian a partir de clulas migenas procedentes de los
55
somitas. La piel de las extremidades en desarrollo tambin es inervada de forma segmentaria. Al principio del desarrollo las ranas primarias ventrales sucesivas se hallan unidas por asas de conexin de fibras nerviosas, especialmente las que inervan a las extremidades (como el plexo humeral). La divisin dorsal de los troncos de estos plexos inervan los msculos extensores y la superficie extensora de la extremidades; las divisiones ventrales de los troncos inervan los msculos flexores y la superficie flexora. Los dermatomas y la inervacin cutnea de las extremidades.
Fig. 5: A, Esquema de un embrin (aproximadamente 41 das) que muestra los miotomas y el sistema muscular en desarrollo. B, Corte transversal del embrin
56
que ilustra los derivados epiaxial e hipoaxial de un miotoma. C, Corte similar de un embrin de siete semanas que muestra las capas musculares formadas a partir del miotoma. 6.2 Pares craneales
A lo largo de las semanas quinta y sexta del desarrollo se forman doce pares de nervios craneales que se clasifican en tres grupos en funcin de su origen embriolgico.
6.3 Pares craneales eferentes somticos
Los nervios pattico (PC IV), motor ocular externo (PC VI), hipogloso (PC XII) y la mayor parte del motor ocular comn (PC III) son homlogos a las races ventrales de los nervios raqudeos (fig. 6). Las clulas de origen de estos nervios se localizan en la columna eferente somtica (derivadas de las placas basales) del tallo enceflico. Sus axones se distribuyen por los msculos derivados de los miotomas de la cabeza (pretico y occipital; figuras).
57
El nervio hipogloso (PC XII) se asemeja a un nervio raqudeo ms que el resto de pares craneales eferentes somticos. El PC XII se forma por la fusin de las fibras de las races ventrales de tres o cuatro nervios occipitales (fig. 6-A). Las races sensitivas correspondientes a las races dorsales de los nervios raqudeos estn ausentes. Las fibras motoras somticas se originan a partir del ncleo hipogloso, formado por clulas motoras similares a las del asta ventral de la medula espinal. Estas fibras dejan la pared ventrolateral del bulbo raqudeo en varios grupos, las races nerviosas hipoglosas, que convergen para formar el tronco comn del PC XII (fig. 6-B). Crecen rostralmente y finalmente inervan los msculos de la lengua, que parecen derivar de miotomas occipitales (fig 7). Al desarrollarse el cuello el nervio hipogloso se sita en una posicin cada vez ms alta.
Fig. 7: Dibujos del sistema muscular en desarrollo. A, Embrin de seis semanas que muestra las regiones de los miotomos de los somitas, que dan lugar a ala mayor parte de los msculos esquelticos. B, Embrin de ocho semanas que muestra la musculatura del tronco y extremidades en desarrollo.
58
El nervio motor ocular externo (PC VI) surge de las clulas nerviosas de las placas basales del metencfalo. Pasa de su superficie ventral a la posterior de los tres miotomas prticos de los cuales se piensa procede el msculo recto externo del ojo.
El nervio pattico (PC IV) procede de clulas nerviosas de la columna eferente somtica de la parte posterior del mecencfalo. Aunque se trata de un nervio motor, sale dorsalmente del tronco enceflico y pasa centralmente hacia el msculo oblicuo superior del ojo.
El nervio motor ocular comn (PC III) inerva la mayora de los msculos del ojo (esto es, rectos superior, inferior e interno y oblicuo inferior) que se cree derivan de los primeros miotomas preticos.
Nervios de los arcos farngeos
Los pares craneales V, VII, IX y X inervan los arcos farngeos embrionarios; por tanto, las estructuras que se desarrollan apartir de estos arcos reciben su innervacin de esos pares craneales (Tabla 10-1).
59
El nervio trigmino (PC V) es el nervio correspondiente al primer arco farngeo, pero posee una rama oftlmica que no es un componente farngeo. El PC V es fundamentalmente sensitivo y constituye principal nervio sensitivo de la cabeza. El ganglio trigmino de gran tamao se encuentra junto al extremo rostral de la protuberancia y sus clulas derivan de la parte ms anterior de la cresta neural. Las prolongaciones centrales de clulas en este ganglio forman la gran raz sensorial del PC V, que penetra en la porcin lateral de la protuberancia. Las prolongaciones perifricas de clulas de este ganglio se separan en tres grandes ramas (nervios oftlmico, maxilar superior y maxilar inferior). Sus fibras sensitivas inervan la piel de la cara, as como el revestimiento de la boca y la nariz (figura). Las fibras motoras del PC V surgen de clulas de la parte mas anterior de la columna eferente visceral especial del metencfalo. El ncleo motor de este nervio se encuentra en la parte media de la protuberancia. Las fibras sales de esta en el lugar de entrada de las fibras sensitivas y pasan a los msculos de la masticacin y a otros que se desarrollan en la prominencia mandibular del primer arco faringes
60
(tabla10-1). El ncleo mesenceflico del PC V se diferencia a partir de clulas del mesencfalo que se extienden rostralmente desde el metencfalo.
El nervio facial (PC VIII) es el nervio del segundo arco farngeo. En su mayor parte esta formado por fibras motoras que surgen principalmente de un grupo nuclear de la columna eferente visceral especial de la porcin caudal de la protuberancia. Estas fibras se distribuyen a los msculos de la expresin facial, as como a otros msculos que se forman en el mesnquima del segundo arco farngeo (tabla 10-1). El pequeo componente eferente visceral general del PC VII termina en los ganglios autnomos perifricos de la cabeza. Las fibras sensitivas del PC VIII surgen de las clulas del ganglio geniculado. Las prolongaciones centrales de estas clulas penetran en la protuberancia, al tiempo que las perifricas pasan al nervio petroso superficial mayor y, atravs de nervio de la cuerda del tmpano, a las papilas gustativas de los dos tercios anteriores de la lengua.
El nervio glosofarngeo (PC IX) es el nervio que corresponde al tercer arco farngeo. Sus fibras motoras surgen de las columna eferentes viscerales especial y, en menos medida, de la columna eferente visceral general de la porcin anterior del mielencfalo. El PC IX se forma apartir de varias raicillas que surgen del bulbo inmediatamente caudales respecto al odo interno en desarrollo. Todas las fibras de la columna eferente visceral especial se distribuyen al msculo estilo farngeo, derivado del mesnquima del tercer arco farngeo (tabla).las fibras eferentes generales se distribuyen al ganglio tico, desde el cual las fibras postganglionares se dirigen a las glndulas partida y linguales posteriores. Las fibras sensitivas del PC IX se distribuyen como fibras aferentes viscerales especiales y sensitivas (fibras gustativas) en la parte posterior de la lengua.
61
El nervio vago (PC X) esta formado por la fusin de los nervios del cuarto y sexto arcos farngeos (tabla 10-1). Posee grandes componentes eferente visceral y aferentes visceral que se distribuyen al corazn, intestino anterior y sus derivados, as como a una extensa porcin del intestino medio. El nervio del cuarto arco farngeo se convierte en el nervio larngeo superior, que inerva el msculo cricotiroideo y los msculos constrictores de la faringe. El nervio correspondiente al sexto arco farngeo se transforma en el nervio larngeo recurrente, que inerva diversos msculos larngeos.
El nervio accesorio (PC XI) tiene dos orgenes distintos (fig. 6). La raz craneal es una extensin posterior del PC X y la raz raqudea sale de los cinco o seis segmentos cervicales craneales de la medula espinal. Las fibras de la raz craneal emergen de la superficie lateral del bulbo, donde se unen al nervio vago e inervan los msculos del paladar blando y msculos intrnsecos de la laringe. Las fibras de la raz raqudea inervan los msculos externocleidomastoideo y trapecio.
6.2 Nervios sensoriales especiales
El nervio olfatorio (PC I) surge del bulbo olfatorio. Las clulas olfatorias son neuronas bipolares que se diferencian apartir de las clulas del revestimiento epitelial del saco nasal primitivo. Los axones de las clulas olfatorias se agrupan en 18 a 20 haces alrededor de los cuales se desarrolla la placa cribiforme del hueso etmoides. Estas fibras sin mielinizar terminan en el bulbo olfatorio.
El nervio tico (PC II) esta formado por ms de un milln de fibras nerviosas que crecen hacia el encfalo aparir de neuroblastos de la retina
62
primaria. Dado que este nervio se desarrolla a partir de la pared evaginada del procencfalo, en realidad representa un trayecto de fibras del encfalo.
El nervio vestibulococlear o auditivo (PC VIII) consta de dos tipos de fibras sensitivas en dos ases; estas fibras de conoces como los nervios vestibular y coclear. El nervio vestibular se origina en los canales semicirculares y el nervio coclear lo hace en el conducto coclear, en el cual se forma el rgano espiral (de Corti). Las neuronas bipolares del nervio vestibular tienen sus cuerpos celulares en el ganglio vestibular. Las prolongaciones centrales de esas clulas terminan en los ncleos vestibulares del suelo del cuarto ventrculo. Los cuerpos celulares de las neuronas bipolares del nervio coclear se encuentra en el ganglio espiral y sus prolongaciones centrales terminan en los ncleos cocleares ventral y dorsal del bulbo.
Desarrollo del sistema nervioso autnomo
A nivel funcional, el sistema autnomo se puede dividir en una porcin simptica (toracolumbar) y otra parasimptica (craneosacra).
Sistema nervioso simptico
Durante la quinta semana, las clulas de la cresta neural de la regin torcica migran a lo largo de cada lado de la medula espinal, en la que forman pares de masas celulares (ganglios) dorso laterales a la aorta (fig. 2). Todos estos ganglios simpticos organizados en segmentos estn conectados en forma de cadena bilateral por fibras nerviosas longitudinales. Estos cordones ganglionares, los
63
troncos simpticos, se localizan a cada lado de los cuerpos vertebrales. Algunas clulas de la cresta neural migran en sentido ventral hacia la aorta y forman neuronas en los ganglios prearticos, como los ganglios celiacos y mesentricos (fig 2). Otras clulas de esta cresta migran hacia el rea del corazn, pulmones y tuvo digestivo, donde forman ganglios terminales en plexos simpticos situados cerca o en el interior de estos rganos.
Cuando se han formado los troncos simpticos, los axones de las neuronas simpticas, localizadas en la columna celular intermedio lateral (asta lateral) de los segmentos lumbares de la medula espinal pasan atravs de la raz ventral de un nervio raqudeo y una rama blanca comunicante (rama de conexin) hasta un ganglio paravertebral (fig. 2). En este lugar pueden establecer una sinapsis con neuronas o bien ascender o descender en el tronco simptico para hacerlo a otros niveles. Otras fibras presinpticas pasan a travs de los ganglios paravertebrales sin hacer sinapsis y forman nervios Esplcnicos para las vsceras. El trayecto de las fibras postsinpticas de realiza a travs de una rama gris comunicante por medio de un ganglio simptico hacia un nervio raqudeo; por consiguiente, los troncos simpticos estn compuestos por fibras ascendentes y descendentes.
7. Sistema nervioso parasimptico Las fibras parasimpticas presinpticas emergen de neuronas de los ncleos del tronco enceflico y de la regin sacra de la medula espinal. Las fibras procedentes del tronco enceflico salen atravs de los nervios motor ocular comn (PC III), facial (PC VII), glosofarngeo (PC IX) y vago (PC X). las neuronas postsinpticas se localizan en los ganglios perifricos o en plexos cercanos o dentro de la estructura inervada (como la pupila del ojo y las glndulas salivales).
64
Referencias bibliogrficas
Gmez de Ferraris. A. Campos Muoz. Histologa y embriologa bucodental. 2 Ed. Editorial Mdica Panamericana 2002. James K. Avery. DJ. Chiego, Jr. Principios de Histologa y embriologa bucal con orientacin clnica. 3 Ed. Editorial Elsevier 2007 Barrancos MJ. Operatoria dental: integracin 4 Ed. Editorial Mdica Panamericana 2006 Graber T, Vanarsdall R, Vig K. Ortodoncia: principios y tcnicas actuales. 4 Ed. Editorial Elsevier 2006 Matamala Vargas F, Fuentes Fernndez R, Ceballos Casanova M. Morfologa y Morfometra del Disco de la Articulacin Temporomandibular en Fetos y Adultos Humanos. Int. J. Morphol., 24(2):245-250, 2006.
Alves N. Study About the Development of the TemporomandibularJoint in the Human Fetuses. Int. J. Morphol.,26(2):309-312, 2008.
65
Вам также может понравиться
- Complejo articular temporomandibular (CATMДокумент37 страницComplejo articular temporomandibular (CATMsandhjОценок пока нет
- Complejo Articular TemporomandibularДокумент14 страницComplejo Articular TemporomandibularJUNIORCHACON100% (28)
- Monografia de ATMДокумент13 страницMonografia de ATMJuanMallmaCartolin40% (5)
- Complejo Articular Témporo MandibularДокумент13 страницComplejo Articular Témporo MandibularGianna ManziОценок пока нет
- Anatomia Funcional y Biomecanica Del Sistema MasticatorioДокумент9 страницAnatomia Funcional y Biomecanica Del Sistema MasticatorioBeatriz Adriana DíazОценок пока нет
- Cap.9 CATM Embriohistología IIДокумент39 страницCap.9 CATM Embriohistología IIPaolaОценок пока нет
- AtmДокумент6 страницAtmWilliam Matias Bobadilla TorresОценок пока нет
- Anatomia Muscular de La AtmДокумент15 страницAnatomia Muscular de La AtmIlsen ManzanillaОценок пока нет
- Informe #2 Generalidades de ATM y Oclusiòn Dental.Документ28 страницInforme #2 Generalidades de ATM y Oclusiòn Dental.Johanna Jaramillo MorenoОценок пока нет
- Alumnos Tema 4 Articulacion TemporomandibuarДокумент38 страницAlumnos Tema 4 Articulacion TemporomandibuarnaddhxielyОценок пока нет
- 2020 Taller de Anatomia Funcional Del Sistema MasticatorioДокумент10 страниц2020 Taller de Anatomia Funcional Del Sistema MasticatorioRaul montes garciaОценок пока нет
- Articulación temporomandibular enДокумент5 страницArticulación temporomandibular enRenata OrueОценок пока нет
- CATMДокумент5 страницCATMenglishhubb1plusunit6.1Оценок пока нет
- Trastornos de La ATM.Документ29 страницTrastornos de La ATM.Tino_21100% (1)
- Articulacion TemporomandibularДокумент12 страницArticulacion TemporomandibularHelbert Tineo SilvaОценок пока нет
- Disco articular ATMДокумент2 страницыDisco articular ATMProfa CortezОценок пока нет
- Anatomia Funcional y Biomecnica Del Sistema Masticatorio VPDFДокумент85 страницAnatomia Funcional y Biomecnica Del Sistema Masticatorio VPDFAlex Estremadoyro67% (3)
- Articulación TemporoДокумент5 страницArticulación TemporosecundariapsolОценок пока нет
- Anatomia de La Articulacion TemporomandibularДокумент7 страницAnatomia de La Articulacion TemporomandibularMirian Jeannine Echeverría Delgado100% (1)
- Articulación TemporomandibularДокумент23 страницыArticulación TemporomandibularGabriel ZunigaОценок пока нет
- Articulación TemporomandibularДокумент23 страницыArticulación TemporomandibularAlejandro RuizОценок пока нет
- CATM anatomía funcionalДокумент36 страницCATM anatomía funcionalKathya DiazОценок пока нет
- Anatomia Funcional y Biomecanica Del Sistema MasticatorioДокумент7 страницAnatomia Funcional y Biomecanica Del Sistema MasticatorioDiego ReyesОценок пока нет
- Apuntes ATMДокумент6 страницApuntes ATMFelipe MaldonadoОценок пока нет
- Sistema EstomatognaticoДокумент4 страницыSistema EstomatognaticoDaniel MolinaОценок пока нет
- Biomecanica Del Atm. Monografia1Документ28 страницBiomecanica Del Atm. Monografia1Alexandra UriarteОценок пока нет
- Tipo Bicondilea Que ExisteДокумент5 страницTipo Bicondilea Que ExisteJulianny Veloz GilОценок пока нет
- seminario oclusion 2Документ7 страницseminario oclusion 2Yusneyly MedinaОценок пока нет
- Atm TrabajoДокумент18 страницAtm TrabajoLaura JaramilloОценок пока нет
- Articulacion Temporomandibular (A.T.M.) : Tpd. Cristian Campos PizarroДокумент17 страницArticulacion Temporomandibular (A.T.M.) : Tpd. Cristian Campos PizarroAida Ines Ugarte EgusquizaОценок пока нет
- Histologia de La ATMДокумент10 страницHistologia de La ATMCarmen Iturriaga Guajardo0% (1)
- Trabajo Oclu Fisiologia MandibularДокумент13 страницTrabajo Oclu Fisiologia MandibularGaymar Daiana Fonseca LinarezОценок пока нет
- Descripción Anatómica de La AtmДокумент4 страницыDescripción Anatómica de La AtmSinai VillacrecesОценок пока нет
- Estructuras Anatomicas Del ATMДокумент7 страницEstructuras Anatomicas Del ATMAlexa Michelle Orozco ContrerasОценок пока нет
- Articulaciones: clasificación, estructura y funcionesДокумент13 страницArticulaciones: clasificación, estructura y funcionesjorge luisОценок пока нет
- ARTICULACION TEMPORO MANDIBULAR - Parte 1Документ23 страницыARTICULACION TEMPORO MANDIBULAR - Parte 1Ariel Armando Mendoza100% (1)
- ATM anatomía 40Документ23 страницыATM anatomía 40Ariel Armando MendozaОценок пока нет
- Tesis Trastornos de La Atm y Su Relaciòn Con La Perdida Dental BiblioДокумент51 страницаTesis Trastornos de La Atm y Su Relaciòn Con La Perdida Dental Bibliojohary chungaОценок пока нет
- Diagnostico Atm MelДокумент49 страницDiagnostico Atm MelFercho HerreraОценок пока нет
- Fisiologia de La Articulacion1Документ5 страницFisiologia de La Articulacion1Alejandra LozaОценок пока нет
- Seminario de ATM y MúsculosДокумент8 страницSeminario de ATM y MúsculosdanielaОценок пока нет
- Atm - OdontologiaДокумент15 страницAtm - OdontologiaGHEILEEMОценок пока нет
- Articulacion TemporomandibularДокумент9 страницArticulacion TemporomandibularMARI JOSE GUZMAN MORALESОценок пока нет
- Reconstruccion de La Articulacion Temporomandibular Con ProtesisДокумент25 страницReconstruccion de La Articulacion Temporomandibular Con ProtesisLina Maria FrancoОценок пока нет
- Oclusión y anatomía funcional de la ATMДокумент69 страницOclusión y anatomía funcional de la ATMSusan Arias100% (1)
- TendonesДокумент7 страницTendonesKennyperalta19Оценок пока нет
- Exposicion Grupo 1 Sistema Estomatognatico 18 04 18Документ50 страницExposicion Grupo 1 Sistema Estomatognatico 18 04 18CesarAmorotoОценок пока нет
- Anatomia BucodentalДокумент21 страницаAnatomia BucodentalFelix Bernabe M. SalazarОценок пока нет
- Desarrollo Del Complejo Articular Temporomandibular EsdmilДокумент6 страницDesarrollo Del Complejo Articular Temporomandibular Esdmilesmil rocaОценок пока нет
- AtmДокумент39 страницAtmELLA NidafeОценок пока нет
- Articulación TemporomandibularДокумент7 страницArticulación TemporomandibularanabelОценок пока нет
- MorfofisiologiaДокумент3 страницыMorfofisiologiavictor riosОценок пока нет
- ATM y Musculos CraneomandibuaresДокумент13 страницATM y Musculos CraneomandibuaresCatalina TrincadoОценок пока нет
- ATM ManualДокумент8 страницATM ManualJorgelina BradaniniОценок пока нет
- Atm EsmilДокумент2 страницыAtm Esmilesmil rocaОценок пока нет
- Anatomia Funcional y Fisiologia Del Sistema MasticatorioДокумент11 страницAnatomia Funcional y Fisiologia Del Sistema Masticatoriojorge alberto bravoОценок пока нет
- Tema 5. AtmДокумент9 страницTema 5. AtmAleisterОценок пока нет
- Espacios Sinoviales y Medios de UniónДокумент4 страницыEspacios Sinoviales y Medios de Unión3nriqueb4rriosОценок пока нет
- Como Tratar Y Prevenir El Dolor De Espalda: Descubre los factores que pueden ocasionarle dolores de espalda a una persona y cuáles son los posibles tratamientos para combatir este malОт EverandComo Tratar Y Prevenir El Dolor De Espalda: Descubre los factores que pueden ocasionarle dolores de espalda a una persona y cuáles son los posibles tratamientos para combatir este malРейтинг: 5 из 5 звезд5/5 (1)
- El Sistema Nervioso CentralДокумент13 страницEl Sistema Nervioso CentralDeisy Jhoana Serna EspitiaОценок пока нет
- Reactivos de NeurofisiologiaДокумент4 страницыReactivos de Neurofisiologiajuana guzmanОценок пока нет
- Ramo de Anatomia IiДокумент66 страницRamo de Anatomia Iimayte.c.glz.05Оценок пока нет
- Enfermedades EspinalesДокумент69 страницEnfermedades EspinalesSuárez DavidОценок пока нет
- Cta 5to Final ExamДокумент5 страницCta 5to Final ExamGisella Marilù Zelaya MendozaОценок пока нет
- Anatomía de La Columna. Escoliosis, Cifosis y LordosisДокумент6 страницAnatomía de La Columna. Escoliosis, Cifosis y LordosispiifjcОценок пока нет
- Qué Es El Encéfalo y Su FunciónesДокумент5 страницQué Es El Encéfalo y Su FunciónesErika isamar Sánchez huamanОценок пока нет
- NeuroanatomíaДокумент25 страницNeuroanatomíaPaulaОценок пока нет
- La Médula Espinal para Tercero de SecundariaДокумент7 страницLa Médula Espinal para Tercero de Secundaria977541897Оценок пока нет
- Mapa ConceptualДокумент1 страницаMapa ConceptualLeonel Zuñiga CristóbalОценок пока нет
- Practica 3 Sistema NerviosoДокумент6 страницPractica 3 Sistema NerviosoZyprexa 17Оценок пока нет
- Médula EspinalДокумент5 страницMédula EspinalYohany Montenegro MuñozОценок пока нет
- MielografíaДокумент19 страницMielografíainbbaamОценок пока нет
- Trauma RaquimedularДокумент8 страницTrauma RaquimedularMarcos TeliasОценок пока нет
- Bases de Psicobiología. El Cerebro Del Caballo Vs El Cerebro Humano. Los Sentidos Del CaballoДокумент6 страницBases de Psicobiología. El Cerebro Del Caballo Vs El Cerebro Humano. Los Sentidos Del CaballoMimi RoedOorОценок пока нет
- La Médula Espinal para Tercero de SecundariaДокумент7 страницLa Médula Espinal para Tercero de Secundariajulio berrocalОценок пока нет
- Adobe Scan 04 Ago. 2023Документ20 страницAdobe Scan 04 Ago. 2023Milanesa RepampanosОценок пока нет
- ANEUROANATOMIAДокумент57 страницANEUROANATOMIAVALENTINA OSORIO RAMIREZОценок пока нет
- Semiología motora y signos meníngeosДокумент23 страницыSemiología motora y signos meníngeosEstrella Llerena100% (1)
- Desarrollo Embrionario de Los Nervios Craneales y RaquídeosДокумент40 страницDesarrollo Embrionario de Los Nervios Craneales y RaquídeosEnrique González50% (2)
- Tareas de NeurologíaДокумент11 страницTareas de NeurologíaRodrigo F. Quiroz SolizОценок пока нет
- Cuál Es La Morfología y Función de La Médula EspinalДокумент3 страницыCuál Es La Morfología y Función de La Médula EspinalFC Aliss100% (1)
- RV 5Документ12 страницRV 5Steven JimenezОценок пока нет
- Estructura Interna Del Tallo CerebralДокумент19 страницEstructura Interna Del Tallo CerebralANGIE NATHALIA GUTIERREZ PARRAОценок пока нет
- Plexopata TraumticaДокумент7 страницPlexopata TraumticaCentro De Radiologia IntervencionistaОценок пока нет
- Los Nervios EspinalesДокумент5 страницLos Nervios EspinalesEduardo Gumeta FarreraОценок пока нет
- Anatomia VestibularДокумент25 страницAnatomia VestibularJoselyn Tamara Rivera RojasОценок пока нет
- Inmovilización Céfalo-Dorso-Lumbar en accidente vehicularДокумент3 страницыInmovilización Céfalo-Dorso-Lumbar en accidente vehicularAntonio JaraОценок пока нет
- Sistema Nervioso Central para Segundo de PrimariaДокумент3 страницыSistema Nervioso Central para Segundo de PrimariaMelina TaxОценок пока нет
- Capitulo 3 Carlson Estructura SNCДокумент62 страницыCapitulo 3 Carlson Estructura SNCXimena Lainfiesta0% (1)