Вам также может понравиться
- Fast Facts: Blastic Plasmacytoid Dendritic Cell Neoplasm: Shedding light on a rare diseaseОт EverandFast Facts: Blastic Plasmacytoid Dendritic Cell Neoplasm: Shedding light on a rare diseaseОценок пока нет
- Abstract BookДокумент90 страницAbstract BookPopescu RoxanaОценок пока нет
- Minimal Residual Disease Testing: Current Innovations and Future DirectionsОт EverandMinimal Residual Disease Testing: Current Innovations and Future DirectionsTodd E. DruleyОценок пока нет
- A Fifth Locus For Otosclerosis, OTSC5, Maps To Chromosome 3q22-24Документ4 страницыA Fifth Locus For Otosclerosis, OTSC5, Maps To Chromosome 3q22-24Ita MaghfirahОценок пока нет
- Article - Genes in AchromatopsiaДокумент8 страницArticle - Genes in AchromatopsiaPablo G. BledtОценок пока нет
- Zufferey. 2012.A 600 KB Deletion Syndrome at 16p11.2 Leads To Energy Imbalance and Neuropsychiatric DisordersДокумент10 страницZufferey. 2012.A 600 KB Deletion Syndrome at 16p11.2 Leads To Energy Imbalance and Neuropsychiatric DisordersEccoОценок пока нет
- Leukodystrophies and Genetic Leukoencephalopathies in Children Specified by Exome Sequencing in An Expanded Gene PanelДокумент9 страницLeukodystrophies and Genetic Leukoencephalopathies in Children Specified by Exome Sequencing in An Expanded Gene PanelEduardo Rios DuboisОценок пока нет
- 2023 Ying Ung Chuin 1680579 Ethesis CompressedДокумент281 страница2023 Ying Ung Chuin 1680579 Ethesis Compressedpilar gabriela hernandez fernandezОценок пока нет
- Gim 201618Документ8 страницGim 201618Ioana PoterasОценок пока нет
- Batten Disease: Features To Facilitate Early Diagnosis: Scientific ReportДокумент6 страницBatten Disease: Features To Facilitate Early Diagnosis: Scientific ReportMaferОценок пока нет
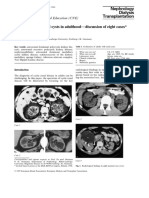
- Polycystic Kidney - Eight CasesДокумент11 страницPolycystic Kidney - Eight CasesKAINAT ANJUMОценок пока нет
- Blastic Plasmacytoid Dendritic Cell NeoplasmeCurrent InsightsДокумент10 страницBlastic Plasmacytoid Dendritic Cell NeoplasmeCurrent InsightsPaúl Otañez MolinaОценок пока нет
- CHAP NO4 Results (Abbas) Rewritten FileДокумент12 страницCHAP NO4 Results (Abbas) Rewritten FileRahid KhanОценок пока нет
- 2006 - JacquemontДокумент8 страниц2006 - JacquemontCarlosОценок пока нет
- SUR - Frei Et Al.Документ6 страницSUR - Frei Et Al.Marcelo EndresОценок пока нет
- SGCE and Myoclonus Dystonia: Motor Characteristics, Diagnostic Criteria and Clinical Predictors of GenotypeДокумент9 страницSGCE and Myoclonus Dystonia: Motor Characteristics, Diagnostic Criteria and Clinical Predictors of GenotypeTalib AdilОценок пока нет
- Medicina 56 00076Документ9 страницMedicina 56 00076Phươngg PhươnggОценок пока нет
- 2008 - StefanssonДокумент6 страниц2008 - StefanssonCarlosОценок пока нет
- Fcell 09 632946Документ12 страницFcell 09 632946charushila patelОценок пока нет
- Aicardi-Goutie'res Syndrome: Neuroradiologic Findings and Follow-UpДокумент6 страницAicardi-Goutie'res Syndrome: Neuroradiologic Findings and Follow-UpclaypotgoldОценок пока нет
- Survey-62-P770 Keratoconus Review - QueratoconoДокумент14 страницSurvey-62-P770 Keratoconus Review - QueratoconoMaría Antonela NicolóОценок пока нет
- Brunklaus (2020) - Biological Concepts in Human Sodium Channel Epilepsies and Their Relevance in Clinical PracticeДокумент13 страницBrunklaus (2020) - Biological Concepts in Human Sodium Channel Epilepsies and Their Relevance in Clinical PracticecicasmileОценок пока нет
- Cat Eye SyndromeДокумент3 страницыCat Eye SyndromeNajia ChoudhuryОценок пока нет
- Yang Cohen Molec IntJDevNeurosci2018Документ6 страницYang Cohen Molec IntJDevNeurosci2018Laura GilОценок пока нет
- The Genetics of Type 2 DiabetesДокумент28 страницThe Genetics of Type 2 DiabetesAmalia Elena BulanceaОценок пока нет
- Fced GenetikДокумент33 страницыFced GenetikyosuanugrahapratamaОценок пока нет
- AMM V 27.07Документ138 страницAMM V 27.07Anonymous 5cHdazK6b100% (1)
- Duplication at Xq13.3-q21.1 With Syndromic Intellectual Disability, A Probable Role For The ATRX GeneДокумент6 страницDuplication at Xq13.3-q21.1 With Syndromic Intellectual Disability, A Probable Role For The ATRX GeneEyyup UctepeОценок пока нет
- Cancer Cells Can Exhibit A Sparing FLASH Effect atДокумент9 страницCancer Cells Can Exhibit A Sparing FLASH Effect atMohamed El-AgamyОценок пока нет
- Genes 12 01513 v2Документ10 страницGenes 12 01513 v2jacopo pruccoliОценок пока нет
- Au Kline HNRNPK-specific DNA MethylationДокумент19 страницAu Kline HNRNPK-specific DNA MethylationjaenciОценок пока нет
- TRIVLI Et Al. 2020. Primary Open Angle Glaucoma Genetics, The Common Variants and Their Clinical Associations (Review)Документ8 страницTRIVLI Et Al. 2020. Primary Open Angle Glaucoma Genetics, The Common Variants and Their Clinical Associations (Review)Thiago RodriguesОценок пока нет
- Skin Health and Disease - 2021 - Zirn - Rothmund Thomson Syndrome Type 1 Caused by Biallelic ANAPC1 Gene MutationsДокумент4 страницыSkin Health and Disease - 2021 - Zirn - Rothmund Thomson Syndrome Type 1 Caused by Biallelic ANAPC1 Gene MutationsEfren BalsecaОценок пока нет
- 4.kharbanda Et Al 2017Документ6 страниц4.kharbanda Et Al 2017MiaОценок пока нет
- EMBO Reports - 2022 - Benfrid - Dengue Virus NS1 Protein Conveys Pro Inflammatory Signals by Docking Onto High DensityДокумент14 страницEMBO Reports - 2022 - Benfrid - Dengue Virus NS1 Protein Conveys Pro Inflammatory Signals by Docking Onto High Densitymatheus.biotechОценок пока нет
- PIIS1098360021012399Документ10 страницPIIS1098360021012399Yenifer Chipana QuispeОценок пока нет
- Articulo de Genetica y Neoplasia Escamosa de La Superficie OcularДокумент14 страницArticulo de Genetica y Neoplasia Escamosa de La Superficie OcularJesús Dávila AlquisirasОценок пока нет
- ContentServer AspДокумент7 страницContentServer AspValentina PereaОценок пока нет
- CNTN6 Mutations Are Risk Factors For Abnormal Auditory Sensory Perception in Autism Spectrum DisordersДокумент9 страницCNTN6 Mutations Are Risk Factors For Abnormal Auditory Sensory Perception in Autism Spectrum DisordersAndres MattarОценок пока нет
- AMM V 27.07Документ138 страницAMM V 27.07paul_calburean7899Оценок пока нет
- Migraña Con Aura: Una Mirada Molecular A Un Problema HereditarioДокумент7 страницMigraña Con Aura: Una Mirada Molecular A Un Problema Hereditariojucrf1218Оценок пока нет
- Molecular Characterization of Melanoma Cases in Denmark Suspected of Genetic PredispositionДокумент16 страницMolecular Characterization of Melanoma Cases in Denmark Suspected of Genetic PredispositionamraovcinaОценок пока нет
- Iae 0000000000001341Документ7 страницIae 0000000000001341fangfang719Оценок пока нет
- Whole Exome Sequencing Revealed Novel Pathogenic Variants in Vietnamese Patients With FEVRДокумент12 страницWhole Exome Sequencing Revealed Novel Pathogenic Variants in Vietnamese Patients With FEVRAmilaОценок пока нет
- Zheng 2013Документ6 страницZheng 2013namiraОценок пока нет
- Epileptic Encephalopathies SYNGAP1Документ7 страницEpileptic Encephalopathies SYNGAP1Saya OtonashiОценок пока нет
- Aku Exon DeletionДокумент6 страницAku Exon DeletionShreyash NaradeОценок пока нет
- 2011 BakirciogluДокумент13 страниц2011 BakirciogluMestrado EducacaoОценок пока нет
- PIIS1098360021012478Документ13 страницPIIS1098360021012478Opale PapaleОценок пока нет
- Multiple Lipomas Linked To An RB1 Gene Mutation in A Large Pedigree With Low Penetrance RetinoblastomaДокумент5 страницMultiple Lipomas Linked To An RB1 Gene Mutation in A Large Pedigree With Low Penetrance RetinoblastomaMaria Burke-ChaussonОценок пока нет
- Congenital Stromal Corneal Dystrophy (2018)Документ12 страницCongenital Stromal Corneal Dystrophy (2018)nadya kharisma amiraОценок пока нет
- Blue EyesДокумент11 страницBlue EyesIlirjan CaneОценок пока нет
- EXOSC8Документ6 страницEXOSC8vuhaipham1407Оценок пока нет
- Genetic Disease Is A Common Cause of Bilateral Childhood Cataract in DenmarkДокумент10 страницGenetic Disease Is A Common Cause of Bilateral Childhood Cataract in DenmarkEvy Alvionita YurnaОценок пока нет
- Congenital Hydrocephalus: Charuta Gavankar Furey, Prince Antwi, and Kristopher Thomas KahleДокумент27 страницCongenital Hydrocephalus: Charuta Gavankar Furey, Prince Antwi, and Kristopher Thomas KahleMerlin MuktialiОценок пока нет
- 05 Molecular Genetics of The EpilepsiesДокумент15 страниц05 Molecular Genetics of The EpilepsiesvenkatesannagarajanОценок пока нет
- Ejhg 201324Документ7 страницEjhg 201324sssssОценок пока нет
- Corneal DystrophyДокумент7 страницCorneal DystrophydyahajengОценок пока нет
- The Immune Microenvironment of HPV-negative OralДокумент8 страницThe Immune Microenvironment of HPV-negative Oraldr israrОценок пока нет
- Winawer Et Al-2018-Annals of NeurologyДокумент14 страницWinawer Et Al-2018-Annals of NeurologyAndoingОценок пока нет
- Biral Etal 2017Документ16 страницBiral Etal 2017bioandreyОценок пока нет
- Generation of External Standards For Real Time PCR (Relative Quantitication)Документ4 страницыGeneration of External Standards For Real Time PCR (Relative Quantitication)Morteza HaghiОценок пока нет
- GMFДокумент2 страницыGMFValery Al-kandariОценок пока нет
- DNA Sequencing: Chemical Method/ Maxam Gilbert MethodДокумент19 страницDNA Sequencing: Chemical Method/ Maxam Gilbert MethodZain YaqoobОценок пока нет
- A Brief History of PCRДокумент54 страницыA Brief History of PCRhabibieОценок пока нет
- C.J. Rocco 2008Документ7 страницC.J. Rocco 2008MariaОценок пока нет
- PCR Based Microbial TypingДокумент76 страницPCR Based Microbial Typingvg04Оценок пока нет
- Methods of IdentificationДокумент54 страницыMethods of IdentificationDhruva Patel0% (1)
- 02 DNA Replication ReadingДокумент1 страница02 DNA Replication Readinglightning TigerОценок пока нет
- (Deja Review) Naheedy, John H. - Orringer, Daniel A - Deja Review. USMLE Step 1-McGraw-Hill Medical (2010)Документ413 страниц(Deja Review) Naheedy, John H. - Orringer, Daniel A - Deja Review. USMLE Step 1-McGraw-Hill Medical (2010)Vansley, V.100% (5)
- Takano 1994Документ6 страницTakano 1994maryjoseОценок пока нет
- U20BTBT01-Biology For EngineersДокумент1 страницаU20BTBT01-Biology For EngineersSiddhi SiddhiОценок пока нет
- Vectors Used in Genetic EngineeringДокумент29 страницVectors Used in Genetic EngineeringkiranshanesОценок пока нет
- 12th Cbse Biology Board Paper 2008 To 2012 SolvedДокумент299 страниц12th Cbse Biology Board Paper 2008 To 2012 Solvedvivekjoseph9578% (18)
- pBR322 MapДокумент1 страницаpBR322 MapCarlos Wesker100% (1)
- BIO 315 Syllabus s2015Документ3 страницыBIO 315 Syllabus s2015JustinОценок пока нет
- Practical 1 GeneticsДокумент20 страницPractical 1 GeneticsizzambrnОценок пока нет
- Training Manual For EmbryoДокумент158 страницTraining Manual For Embryotolimania2951Оценок пока нет
- A HandsДокумент13 страницA HandsVeyari RamirezОценок пока нет
- Andrew Labenek Abortion CompleteДокумент12 страницAndrew Labenek Abortion Completeapi-354532409Оценок пока нет
- Genetic Calculator (Budgerigar) Help File: © 2016 K YorkeДокумент22 страницыGenetic Calculator (Budgerigar) Help File: © 2016 K YorkeMarlindaОценок пока нет
- Translation NotesДокумент4 страницыTranslation NotesRishikesh BhintadeОценок пока нет
- Gene PredictionДокумент50 страницGene Predictionsaeedahmad901Оценок пока нет
- Science 8 Week 3 Quarter 4Документ9 страницScience 8 Week 3 Quarter 4Juliana ChuaОценок пока нет
- Lifespan Development Lives in Context 1st Edition Kuther Test BankДокумент24 страницыLifespan Development Lives in Context 1st Edition Kuther Test BankDawnCalderonwmap100% (34)
- Thrushfield Chap2Документ21 страницаThrushfield Chap2Βενιαμίν στέμμαОценок пока нет
- Ms Campbell Protein Synthesis Practice Questions Regents LeДокумент6 страницMs Campbell Protein Synthesis Practice Questions Regents Lekim del mundoОценок пока нет
- KLP 6 Berangkai Dan Pindah Silang GENETIKAДокумент11 страницKLP 6 Berangkai Dan Pindah Silang GENETIKASakina Ayu PutriОценок пока нет
- Poodle Color GeneticsДокумент10 страницPoodle Color GeneticsGerardo Martell MartínezОценок пока нет
- Assignment On Animal Cloning Edexcel Igcse Biology 9-1Документ8 страницAssignment On Animal Cloning Edexcel Igcse Biology 9-1Rawdatul JannahОценок пока нет
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsОт EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsОценок пока нет
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisОт EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisРейтинг: 4.5 из 5 звезд4.5/5 (42)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeОт EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeРейтинг: 2 из 5 звезд2/5 (1)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsОт EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsРейтинг: 5 из 5 звезд5/5 (1)
- The Age of Magical Overthinking: Notes on Modern IrrationalityОт EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityРейтинг: 4 из 5 звезд4/5 (24)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedОт EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedРейтинг: 5 из 5 звезд5/5 (80)
- The Obesity Code: Unlocking the Secrets of Weight LossОт EverandThe Obesity Code: Unlocking the Secrets of Weight LossРейтинг: 4 из 5 звезд4/5 (6)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaОт EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsОт EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsРейтинг: 4.5 из 5 звезд4.5/5 (169)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDОт EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDРейтинг: 5 из 5 звезд5/5 (1)
- Why We Die: The New Science of Aging and the Quest for ImmortalityОт EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityРейтинг: 4 из 5 звезд4/5 (3)
- Gut: the new and revised Sunday Times bestsellerОт EverandGut: the new and revised Sunday Times bestsellerРейтинг: 4 из 5 звезд4/5 (392)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeОт EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeРейтинг: 4.5 из 5 звезд4.5/5 (253)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsОт EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsРейтинг: 3.5 из 5 звезд3.5/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisОт EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisРейтинг: 3.5 из 5 звезд3.5/5 (2)
- Sleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningОт EverandSleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningРейтинг: 4 из 5 звезд4/5 (3)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.От EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Рейтинг: 4.5 из 5 звезд4.5/5 (110)
- To Explain the World: The Discovery of Modern ScienceОт EverandTo Explain the World: The Discovery of Modern ScienceРейтинг: 3.5 из 5 звезд3.5/5 (51)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessОт EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessРейтинг: 4.5 из 5 звезд4.5/5 (328)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisОт EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisРейтинг: 4 из 5 звезд4/5 (1)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryОт EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryРейтинг: 4 из 5 звезд4/5 (44)
- Summary: The Myth of Normal: Trauma, Illness, and Healing in a Toxic Culture By Gabor Maté MD & Daniel Maté: Key Takeaways, Summary & AnalysisОт EverandSummary: The Myth of Normal: Trauma, Illness, and Healing in a Toxic Culture By Gabor Maté MD & Daniel Maté: Key Takeaways, Summary & AnalysisРейтинг: 4 из 5 звезд4/5 (9)