Вам также может понравиться
- Open Paw MatrixДокумент6 страницOpen Paw MatrixShoshannah Forbes100% (2)
- Exposure Therapy: Rethinking the Model - Refining the MethodОт EverandExposure Therapy: Rethinking the Model - Refining the MethodPeter NeudeckОценок пока нет
- Liotti 2006. A Model of Dissociation Based On Attachment Theory and ResearchДокумент20 страницLiotti 2006. A Model of Dissociation Based On Attachment Theory and Researchzzmarianozz100% (1)
- Microfinance Ass 1Документ15 страницMicrofinance Ass 1Willard MusengeyiОценок пока нет
- The First Voyage Round The World by MageДокумент405 страницThe First Voyage Round The World by MageGift Marieneth LopezОценок пока нет
- FDFHFDHДокумент33 страницыFDFHFDHAnonymous KrfJpXb4iNОценок пока нет
- Craske, Treanor Et Al. 2014Документ14 страницCraske, Treanor Et Al. 2014John NathanОценок пока нет
- BJP 17 0241+final+Документ45 страницBJP 17 0241+final+rainbowee2004Оценок пока нет
- Fear Generalization in Humans-Impact of Verbal InstructionsДокумент6 страницFear Generalization in Humans-Impact of Verbal Instructionsasrapx100% (1)
- Social Beliefs and Judgements Reading 2Документ32 страницыSocial Beliefs and Judgements Reading 2joms magОценок пока нет
- Wells.1995.Social Phobia: The Role of In-Situation Safety Behaviors in Maintaining Anxiety and Negative BeliefsДокумент9 страницWells.1995.Social Phobia: The Role of In-Situation Safety Behaviors in Maintaining Anxiety and Negative BeliefsGanellОценок пока нет
- Journal Pre-Proof: Behaviour Research and TherapyДокумент177 страницJournal Pre-Proof: Behaviour Research and TherapyTomas JosephОценок пока нет
- RESEARCHMETHODSДокумент32 страницыRESEARCHMETHODSAqib ArshadОценок пока нет
- Characteristics of Testable HypothesesДокумент30 страницCharacteristics of Testable HypothesesMarivic Diano67% (3)
- Barry 2016Документ12 страницBarry 2016José VegaОценок пока нет
- Mertens 2018Документ16 страницMertens 2018jorge9000000Оценок пока нет
- FearLearning AldanaAlegríaДокумент3 страницыFearLearning AldanaAlegríaAldana AlegríaОценок пока нет
- Green, Ricky and Douglas, Karen (2018) Anxious Attachment and Belief in Conspiracy TheoriesДокумент38 страницGreen, Ricky and Douglas, Karen (2018) Anxious Attachment and Belief in Conspiracy Theoriesjc2271Оценок пока нет
- A Meta-Analysis of Conditioned Fear Generalization in Anxiety-Related DisordersДокумент10 страницA Meta-Analysis of Conditioned Fear Generalization in Anxiety-Related DisordersGabriel Dos SantosОценок пока нет
- Personality and Reinforcement in AssociaДокумент25 страницPersonality and Reinforcement in AssociaAnggaОценок пока нет
- Sychological Esearch: 02/14 - TimmyДокумент8 страницSychological Esearch: 02/14 - TimmymysterioushumaneОценок пока нет
- Cognitive Mechanisms Underlying Lying To Questions: Response Time As A Cue To DeceptionДокумент20 страницCognitive Mechanisms Underlying Lying To Questions: Response Time As A Cue To DeceptionSergio Blanes CáceresОценок пока нет
- Between Vulnerability and Resilience: A Network Analysis of Fluctuations in Cognitive Risk and Protective Factors Following Remission From DepressionДокумент37 страницBetween Vulnerability and Resilience: A Network Analysis of Fluctuations in Cognitive Risk and Protective Factors Following Remission From DepressionGILMAR DA SILVA JERONIMOОценок пока нет
- Evaluating ResearchДокумент3 страницыEvaluating ResearchCwen Jazlyn SumalinogОценок пока нет
- Self-Esteem, Self-Affirmation, and Schadenfreude: Brief ReportДокумент5 страницSelf-Esteem, Self-Affirmation, and Schadenfreude: Brief ReportCatarina C.Оценок пока нет
- The Influence of Improper Sets of Information On Judgment: How Irrelevant Information Can Bias Judged ProbabilityДокумент20 страницThe Influence of Improper Sets of Information On Judgment: How Irrelevant Information Can Bias Judged ProbabilityJesse PanchoОценок пока нет
- Clark Anx DisordДокумент23 страницыClark Anx Disordeschermer2658Оценок пока нет
- Competition Superstition and The Illusion of ControlДокумент17 страницCompetition Superstition and The Illusion of ControlIlda Sacirovic AdemovicОценок пока нет
- 1.defense Mechanism Is Predicted by AttachmentДокумент9 страниц1.defense Mechanism Is Predicted by AttachmentRolazski123Оценок пока нет
- Blavatsky y 2009Документ11 страницBlavatsky y 2009Amir HayatОценок пока нет
- STATISTIC 11891 - Chapter - 5Документ34 страницыSTATISTIC 11891 - Chapter - 5steveОценок пока нет
- Amos Tversky and Daniel Kahneman Probabilistic ReasoningДокумент26 страницAmos Tversky and Daniel Kahneman Probabilistic ReasoningNiky PapucОценок пока нет
- 1989.2. Beliefs and Inferences-A Test of A Rational-Emotive Hypothesi 2 (Windy Dryden)Документ9 страниц1989.2. Beliefs and Inferences-A Test of A Rational-Emotive Hypothesi 2 (Windy Dryden)fiva.vcОценок пока нет
- Ratcliff Smith 04Документ35 страницRatcliff Smith 04Jay GastbyОценок пока нет
- J Neuropharm 2011 02 023Документ10 страницJ Neuropharm 2011 02 023Isabely AmabilyОценок пока нет
- Attachment Trauma 2011 2Документ35 страницAttachment Trauma 2011 2Sónia Mendes Barbosa100% (1)
- Semcho Et Al 2022Документ43 страницыSemcho Et Al 2022Diana SarcaОценок пока нет
- Hernandez, Preston - 2013 - Disfluency Disrupts The Confirmation BiasДокумент5 страницHernandez, Preston - 2013 - Disfluency Disrupts The Confirmation Biasnicolas mossoОценок пока нет
- Optimism SuperstitionДокумент10 страницOptimism SuperstitionChaps Bryan CatamaОценок пока нет
- AnxStut - JFD - 2004 - Social Anxiety in Stuttering - Measuring Negative Social ExpectanciesДокумент12 страницAnxStut - JFD - 2004 - Social Anxiety in Stuttering - Measuring Negative Social ExpectanciesfpОценок пока нет
- Relative Salience Versus Relative Validity: Cue Salience Influences Blocking in Human Associative LearningДокумент17 страницRelative Salience Versus Relative Validity: Cue Salience Influences Blocking in Human Associative LearningotiliaglamОценок пока нет
- Bengson, Experimental Attacks On Intuitions and AnswersДокумент38 страницBengson, Experimental Attacks On Intuitions and AnswersTiaraju Molina AndreazzaОценок пока нет
- Temper PrecursorsДокумент13 страницTemper PrecursorsJoeОценок пока нет
- Grupe-Nitschke Anxiety 2013Документ14 страницGrupe-Nitschke Anxiety 2013pufpuruf1976100% (1)
- 1-Extinción Vicaria - 2013Документ10 страниц1-Extinción Vicaria - 2013Irene C. CruzОценок пока нет
- Escala ResilienciaДокумент28 страницEscala Resilienciacrista776Оценок пока нет
- An Empirical Test of The Three-Step Theory of Suicide in U.K. University StudentsДокумент10 страницAn Empirical Test of The Three-Step Theory of Suicide in U.K. University StudentsDavidОценок пока нет
- Exemplification Effects in The Promotion of Safety and HealthДокумент19 страницExemplification Effects in The Promotion of Safety and Healthdesiree_markham100Оценок пока нет
- Best AnswerДокумент167 страницBest AnswerAlex DarmawanОценок пока нет
- Sex Differences Are Not Attenuated byДокумент40 страницSex Differences Are Not Attenuated byNolber C ChОценок пока нет
- The Critical Role of Group Studies in NeuropsychologyДокумент14 страницThe Critical Role of Group Studies in NeuropsychologymatttОценок пока нет
- Attachment Dispositions and Human Defensive Behavior 2014 Personality and Individual DifferencesДокумент5 страницAttachment Dispositions and Human Defensive Behavior 2014 Personality and Individual DifferencesArielОценок пока нет
- A Cognitive Model of Persecutory DelusionsДокумент17 страницA Cognitive Model of Persecutory DelusionsSabrina MorelliОценок пока нет
- Validity IssuesДокумент4 страницыValidity IssuesCUCU KURNIASIHОценок пока нет
- Overwalle 1992Документ17 страницOverwalle 1992vladimir216Оценок пока нет
- Final Revision For Pi 9.23.17Документ52 страницыFinal Revision For Pi 9.23.17Ahmed Awan AhmedОценок пока нет
- Running Head: Conditioning Theory and The Development of AnxietyДокумент53 страницыRunning Head: Conditioning Theory and The Development of AnxietyABCОценок пока нет
- Defending One S Worldview UndeДокумент13 страницDefending One S Worldview UndecutkilerОценок пока нет
- Buer Linton 2002 Cita 10Документ7 страницBuer Linton 2002 Cita 10Claudia Andrea Bahamondes WeissohnОценок пока нет
- Effective: and Ineffective Use of Fear inДокумент6 страницEffective: and Ineffective Use of Fear intrisnaОценок пока нет
- Olson Et Al 2011Документ25 страницOlson Et Al 2011Alina RotaruОценок пока нет
- What Do I Do Now - Intolerance of Uncertainty Is Associated With Discrete Patterns of Anticipatory Physiological Responding To Different Uncertain ContextsДокумент49 страницWhat Do I Do Now - Intolerance of Uncertainty Is Associated With Discrete Patterns of Anticipatory Physiological Responding To Different Uncertain ContextsMIGUEL ANGEL JIMENEZ HUERTASОценок пока нет
- Defense Mechanisms: Theoretical, Research and Clinical PerspectivesОт EverandDefense Mechanisms: Theoretical, Research and Clinical PerspectivesРейтинг: 4 из 5 звезд4/5 (1)
- Myths of High ProteinДокумент6 страницMyths of High ProteinShoshannah ForbesОценок пока нет
- ABCsof Dog LifeДокумент199 страницABCsof Dog LifeShoshannah Forbes100% (1)
- Level and 2 TestsДокумент2 страницыLevel and 2 TestsShoshannah ForbesОценок пока нет
- Breeds and Behavior - Bradley Reprint From Bark NEWДокумент3 страницыBreeds and Behavior - Bradley Reprint From Bark NEWShoshannah ForbesОценок пока нет
- Myths of High ProteinДокумент6 страницMyths of High ProteinShoshannah ForbesОценок пока нет
- Cotw Handouts 2011Документ8 страницCotw Handouts 2011Shoshannah ForbesОценок пока нет
- Engineered Competitive Swimsuits - Part 1Документ2 страницыEngineered Competitive Swimsuits - Part 1Vasant KothariОценок пока нет
- Efficacy of A Combination of Imidacloprid 10% - Moxidectin 2Документ9 страницEfficacy of A Combination of Imidacloprid 10% - Moxidectin 2Shoshannah ForbesОценок пока нет
- Personality Traits in The Domestic Dog (Canis Familiaris)Документ23 страницыPersonality Traits in The Domestic Dog (Canis Familiaris)Shoshannah ForbesОценок пока нет
- Proshow MacДокумент29 страницProshow MacShoshannah ForbesОценок пока нет
- HSUS-Purebred Paradox PDFДокумент8 страницHSUS-Purebred Paradox PDFShoshannah ForbesОценок пока нет
- NAIA - The Evolution of Modern-Day Dog TrainingДокумент9 страницNAIA - The Evolution of Modern-Day Dog TrainingShoshannah ForbesОценок пока нет
- PED RebuttalДокумент5 страницPED RebuttalS. Raelynne BecerraОценок пока нет
- Shelter ProgramДокумент42 страницыShelter ProgramfcalaguasОценок пока нет
- The New Definition of "Good Dog" - Ruffly SpeakingRuffly SpeakingДокумент20 страницThe New Definition of "Good Dog" - Ruffly SpeakingRuffly SpeakingShoshannah ForbesОценок пока нет
- J VET Diagn Invest 2001 La Perle 252 5Документ5 страницJ VET Diagn Invest 2001 La Perle 252 5Shoshannah ForbesОценок пока нет
- Dog Obedience Training Tips For Dog Club MembersДокумент3 страницыDog Obedience Training Tips For Dog Club MembersShoshannah ForbesОценок пока нет
- NAIA - The Evolution of Modern-Day Dog TrainingДокумент9 страницNAIA - The Evolution of Modern-Day Dog TrainingShoshannah ForbesОценок пока нет
- WF 81 BarДокумент5 страницWF 81 BarShoshannah ForbesОценок пока нет
- The Problem With ShockДокумент3 страницыThe Problem With ShockShoshannah ForbesОценок пока нет
- Snbehaviorbonedatasnapshot 3Документ4 страницыSnbehaviorbonedatasnapshot 3api-257186144Оценок пока нет
- Factors Linked To Territorial Aggression in DogsДокумент5 страницFactors Linked To Territorial Aggression in DogsShoshannah ForbesОценок пока нет
- Michael Jack 1975 - Positive Negative Reinforcement A Distinction That Is No Longer Necessary or A Better WДокумент13 страницMichael Jack 1975 - Positive Negative Reinforcement A Distinction That Is No Longer Necessary or A Better WShoshannah ForbesОценок пока нет
- Premack PrincipleДокумент6 страницPremack PrincipleShoshannah ForbesОценок пока нет
- V63N02 087Документ5 страницV63N02 087Shoshannah ForbesОценок пока нет
- Alexander ThesisДокумент96 страницAlexander ThesisShoshannah ForbesОценок пока нет
- Michael Jack 1975 - Positive Negative Reinforcement A Distinction That Is No Longer Necessary or A Better WДокумент13 страницMichael Jack 1975 - Positive Negative Reinforcement A Distinction That Is No Longer Necessary or A Better WShoshannah ForbesОценок пока нет
- ThesisДокумент52 страницыThesisShoshannah ForbesОценок пока нет
- Lesson 1 Q3 Figure Life DrawingДокумент10 страницLesson 1 Q3 Figure Life DrawingCAHAPОценок пока нет
- Brosur YSIO X.preeДокумент20 страницBrosur YSIO X.preeRadiologi RSUD KilisuciОценок пока нет
- Barrett Beyond Psychometrics 2003 AugmentedДокумент34 страницыBarrett Beyond Psychometrics 2003 AugmentedRoy Umaña CarrilloОценок пока нет
- 9881 enДокумент345 страниц9881 enSaid BenОценок пока нет
- GLOBE2Документ7 страницGLOBE2mba departmentОценок пока нет
- Directorate of Technical Education, Admission Committee For Professional Courses (ACPC), GujaratДокумент2 страницыDirectorate of Technical Education, Admission Committee For Professional Courses (ACPC), GujaratgamailkabaaaapОценок пока нет
- Paul Spicker - The Welfare State A General TheoryДокумент162 страницыPaul Spicker - The Welfare State A General TheoryTista ArumОценок пока нет
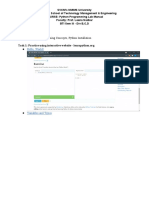
- Week - 2 Lab - 1 - Part I Lab Aim: Basic Programming Concepts, Python InstallationДокумент13 страницWeek - 2 Lab - 1 - Part I Lab Aim: Basic Programming Concepts, Python InstallationSahil Shah100% (1)
- L5V 00004Документ2 страницыL5V 00004Jhon LinkОценок пока нет
- Lancru hzj105 DieselДокумент2 страницыLancru hzj105 DieselMuhammad MasdukiОценок пока нет
- Akilas Mebrahtom ResumeДокумент1 страницаAkilas Mebrahtom ResumeAnonymous lLpBfQОценок пока нет
- Cambridge IGCSE: CHEMISTRY 0620/42Документ12 страницCambridge IGCSE: CHEMISTRY 0620/42Khairun nissaОценок пока нет
- Chapter 2.3.3 History of Visual Arts Modernism Post ModernismДокумент17 страницChapter 2.3.3 History of Visual Arts Modernism Post ModernismKim Ashley SarmientoОценок пока нет
- Parallel Port Programming With DelphiДокумент4 страницыParallel Port Programming With Delphiramadhan1933Оценок пока нет
- Preparing For CPHQ .. An Overview of Concepts: Ghada Al-BarakatiДокумент109 страницPreparing For CPHQ .. An Overview of Concepts: Ghada Al-BarakatiBilal SalamehОценок пока нет
- CAREДокумент11 страницCARELuis SementeОценок пока нет
- A2Документ4 страницыA2Akshay KumarОценок пока нет
- SAFE RC Design ForДокумент425 страницSAFE RC Design ForMarlon Braggian Burgos FloresОценок пока нет
- GCP Vol 2 PDF (2022 Edition)Документ548 страницGCP Vol 2 PDF (2022 Edition)Sergio AlvaradoОценок пока нет
- Ismb ItpДокумент3 страницыIsmb ItpKumar AbhishekОценок пока нет
- Prognostic Factors and Management of Patients With Choanal AtresiaДокумент7 страницPrognostic Factors and Management of Patients With Choanal Atresiafarah maulida martaОценок пока нет
- Coal Bottom Ash As Sand Replacement in ConcreteДокумент9 страницCoal Bottom Ash As Sand Replacement in ConcretexxqОценок пока нет
- Principles of Supply Chain Management A Balanced Approach 4th Edition Wisner Solutions ManualДокумент36 страницPrinciples of Supply Chain Management A Balanced Approach 4th Edition Wisner Solutions Manualoutlying.pedantry.85yc100% (28)
- Bubble Test Direct Pressure InserviceДокумент3 страницыBubble Test Direct Pressure InserviceEdAlmОценок пока нет
- Acetylcysteine 200mg (Siran, Reolin)Документ5 страницAcetylcysteine 200mg (Siran, Reolin)ddandan_2Оценок пока нет
- Lab Manual Switchgear and Protection SapДокумент46 страницLab Manual Switchgear and Protection SapYash MaheshwariОценок пока нет
- 2022 Tony Elumelu EntrepreneursДокумент10 страниц2022 Tony Elumelu EntrepreneursOladosu EmmanuelОценок пока нет
- Introduction To Password Cracking Part 1Документ8 страницIntroduction To Password Cracking Part 1Tibyan MuhammedОценок пока нет