Вам также может понравиться
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Classifications of AssessmentsДокумент11 страницClassifications of AssessmentsClaire CatapangОценок пока нет
- Working Capital in YamahaДокумент64 страницыWorking Capital in YamahaRenu Jindal50% (2)
- Work ProblemsДокумент19 страницWork ProblemsOfelia DavidОценок пока нет
- Wilcoxon Matched Pairs Signed Rank TestДокумент3 страницыWilcoxon Matched Pairs Signed Rank TestDawn Ilish Nicole DiezОценок пока нет
- Recommendations For Students With High Functioning AutismДокумент7 страницRecommendations For Students With High Functioning AutismLucia SaizОценок пока нет
- Damodaram Sanjivayya National Law University Visakhapatnam, A.P., IndiaДокумент25 страницDamodaram Sanjivayya National Law University Visakhapatnam, A.P., IndiaSumanth RoxtaОценок пока нет
- Networker Performance Tuning PDFДокумент49 страницNetworker Performance Tuning PDFHarry SharmaОценок пока нет
- IMS Objectives Targets 2Документ2 страницыIMS Objectives Targets 2FaridUddin Ahmed100% (3)
- Assembler Pass 2Документ5 страницAssembler Pass 2AnuОценок пока нет
- Farmer Producer Companies in OdishaДокумент34 страницыFarmer Producer Companies in OdishaSuraj GantayatОценок пока нет
- Costbenefit Analysis 2015Документ459 страницCostbenefit Analysis 2015TRÂM NGUYỄN THỊ BÍCHОценок пока нет
- 18 June 2020 12:03: New Section 1 Page 1Документ4 страницы18 June 2020 12:03: New Section 1 Page 1KarthikNayakaОценок пока нет
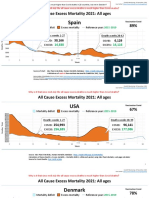
- Countries EXCESS DEATHS All Ages - 15nov2021Документ21 страницаCountries EXCESS DEATHS All Ages - 15nov2021robaksОценок пока нет
- CT SizingДокумент62 страницыCT SizingMohamed TalebОценок пока нет
- CX Programmer Operation ManualДокумент536 страницCX Programmer Operation ManualVefik KaraegeОценок пока нет
- TCL LD24D50 - Chassis MS09A-LA - (TKLE2413D) - Manual de Servicio PDFДокумент41 страницаTCL LD24D50 - Chassis MS09A-LA - (TKLE2413D) - Manual de Servicio PDFFabian OrtuzarОценок пока нет
- Ron Kangas - IoanДокумент11 страницRon Kangas - IoanBogdan SoptereanОценок пока нет
- UTP Student Industrial ReportДокумент50 страницUTP Student Industrial ReportAnwar HalimОценок пока нет
- Outdoor Air Pollution: Sources, Health Effects and SolutionsДокумент20 страницOutdoor Air Pollution: Sources, Health Effects and SolutionsCamelia RadulescuОценок пока нет
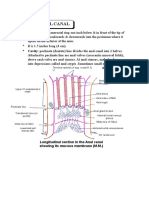
- Anatomy Anal CanalДокумент14 страницAnatomy Anal CanalBela Ronaldoe100% (1)
- Checklist & Guideline ISO 22000Документ14 страницChecklist & Guideline ISO 22000Documentos Tecnicos75% (4)
- School of Mathematics 2021 Semester 1 MAT1841 Continuous Mathematics For Computer Science Assignment 1Документ2 страницыSchool of Mathematics 2021 Semester 1 MAT1841 Continuous Mathematics For Computer Science Assignment 1STEM Education Vung TauОценок пока нет
- Chemistry: Crash Course For JEE Main 2020Документ18 страницChemistry: Crash Course For JEE Main 2020Sanjeeb KumarОценок пока нет
- C C C C: "P P P P PДокумент25 страницC C C C: "P P P P PShalu Dua KatyalОценок пока нет
- Principals' Leadership Styles and Student Academic Performance in Secondary Schools in Ekiti State, NigeriaДокумент12 страницPrincipals' Leadership Styles and Student Academic Performance in Secondary Schools in Ekiti State, NigeriaiqraОценок пока нет
- Module 1: Overview of Applied Behaviour Analysis (ABA)Документ37 страницModule 1: Overview of Applied Behaviour Analysis (ABA)PriyaОценок пока нет
- Carriage RequirementsДокумент63 страницыCarriage RequirementsFred GrosfilerОценок пока нет
- Article An Incident and Injury Free Culture Changing The Face of Project Operations Terra117 2Документ6 страницArticle An Incident and Injury Free Culture Changing The Face of Project Operations Terra117 2nguyenthanhtuan_ecoОценок пока нет
- Powerpoint Speaker NotesДокумент4 страницыPowerpoint Speaker Notesapi-273554555Оценок пока нет
- Project ManagementДокумент11 страницProject ManagementBonaventure NzeyimanaОценок пока нет