Вам также может понравиться
- Mullins 1992Документ6 страницMullins 1992yalocim666Оценок пока нет
- Brief CommunicationДокумент6 страницBrief CommunicationFranz BackenbauerОценок пока нет
- Gastrointestinal Morphology and Absorption of Monosaccharides in Conditioned To Different Types and Levels of Dietary FibreДокумент13 страницGastrointestinal Morphology and Absorption of Monosaccharides in Conditioned To Different Types and Levels of Dietary FibreayakashimajorinОценок пока нет
- Anthocyanins Induce Cholesterol Efflux From Mouse Peritoneal MacrophagesДокумент10 страницAnthocyanins Induce Cholesterol Efflux From Mouse Peritoneal MacrophagesTri Anna FitrianiОценок пока нет
- Bioquímica Experimentó en RatasДокумент6 страницBioquímica Experimentó en RatasJuan David DuranОценок пока нет
- 1 s2.0 S002203021100213X MainДокумент9 страниц1 s2.0 S002203021100213X MainJuniClaudia13Оценок пока нет
- Microsome For HorakДокумент4 страницыMicrosome For HorakCarolina Torres PalazzoloОценок пока нет
- FoodChem 2018 JadanДокумент28 страницFoodChem 2018 JadanGaby Garcia GuerreroОценок пока нет
- Nutrition: Basic Nutritional InvestigationДокумент8 страницNutrition: Basic Nutritional InvestigationkasabeОценок пока нет
- Ensilaje de PescadoДокумент7 страницEnsilaje de PescadojjrochismОценок пока нет
- Dr. Blanco ArticleДокумент11 страницDr. Blanco ArticleCarlosОценок пока нет
- 2010 - Ana P. Martins Et Al - Conjugated Linoleic Acid Reduces Permeability and Fluidity of Adipose Plasma Membranes From Obese Zucker RatsДокумент6 страниц2010 - Ana P. Martins Et Al - Conjugated Linoleic Acid Reduces Permeability and Fluidity of Adipose Plasma Membranes From Obese Zucker RatsVnus_das_pelesОценок пока нет
- Fructans of Jerusalem ArtichokesДокумент7 страницFructans of Jerusalem ArtichokesAsmaa OtifyОценок пока нет
- How Noscapine Metabolise Heme?Документ15 страницHow Noscapine Metabolise Heme?Chief editorОценок пока нет
- International Journal of Biological MacromoleculesДокумент5 страницInternational Journal of Biological MacromoleculesYohana MelianthaОценок пока нет
- Glycolysis-Deficient: Studies Cells That Production of TumorДокумент5 страницGlycolysis-Deficient: Studies Cells That Production of TumorFabian BalderrabanoОценок пока нет
- 01 1159.fullДокумент12 страниц01 1159.fullCiandeb GracianoОценок пока нет
- Effect of Sodium Metabisulfite On Lipid Peroxidation and Enzyme Activi-Ties in Adult Rat Stomach and SpleenДокумент8 страницEffect of Sodium Metabisulfite On Lipid Peroxidation and Enzyme Activi-Ties in Adult Rat Stomach and Spleenام محمدОценок пока нет
- Effect of Wakame On Non Alcoholic Fatty LiverДокумент14 страницEffect of Wakame On Non Alcoholic Fatty LiverCătălina RădulescuОценок пока нет
- Significance of Nutrient Digestibility in Horse Nutrition - A ReviewДокумент19 страницSignificance of Nutrient Digestibility in Horse Nutrition - A ReviewTóthi RóbertОценок пока нет
- 1 s2.0 S2405580821001813 MainДокумент8 страниц1 s2.0 S2405580821001813 MainkhudhoriОценок пока нет
- Dietary Choline Supplementation On Growth PerformanceДокумент8 страницDietary Choline Supplementation On Growth PerformanceIsarosaeОценок пока нет
- Malabsorption GuideДокумент55 страницMalabsorption Guidejimboy1884Оценок пока нет
- Seminario 4A Regulacion de La GlicemiaДокумент6 страницSeminario 4A Regulacion de La GlicemiaEdol LopezОценок пока нет
- Zhang - 2017 - Quantification of Lactate Isotopologues and IsotopomersДокумент12 страницZhang - 2017 - Quantification of Lactate Isotopologues and IsotopomersMokr Guerrero EsperanzaОценок пока нет
- Food ChemistryДокумент5 страницFood ChemistryLa Ode Muhammad FitrawanОценок пока нет
- Digestion Absorption 12 2019Документ14 страницDigestion Absorption 12 2019Karyan ShinОценок пока нет
- Chem. Senses 32: 41-49, 2007Документ9 страницChem. Senses 32: 41-49, 2007gas_yoe2Оценок пока нет
- Peta KonsepДокумент7 страницPeta KonsepNaura HasnaОценок пока нет
- Metabolic Analysis of BALB/c Mice (Mus Musculus) Exposed in High Fat and Protein Diet Using Human Diagnostics Test KitsДокумент16 страницMetabolic Analysis of BALB/c Mice (Mus Musculus) Exposed in High Fat and Protein Diet Using Human Diagnostics Test KitsEmotholy LogicalОценок пока нет
- colarma,+WRS 28-4 12161Документ11 страницcolarma,+WRS 28-4 12161Luidi MarquesОценок пока нет
- Huang 2013Документ10 страницHuang 2013Stefania DavidОценок пока нет
- Sterol Carrier Protein and Fatty Acid-Binding ProteinДокумент7 страницSterol Carrier Protein and Fatty Acid-Binding ProteinMartin OreОценок пока нет
- Gut's Role in Lipid HomeostasisДокумент25 страницGut's Role in Lipid HomeostasisRessy GuslitaОценок пока нет
- Ostion Impact of Two Commercial in Vivo TransportДокумент6 страницOstion Impact of Two Commercial in Vivo TransportNathaly Montoya CamachoОценок пока нет
- Myo-Inositol Deficiency: Studies On The Mechanism of Lactation-Dependent Fatty Liver Formation in The Rat1Документ9 страницMyo-Inositol Deficiency: Studies On The Mechanism of Lactation-Dependent Fatty Liver Formation in The Rat1Hitesh VermaОценок пока нет
- Metformina 1992 Nao HiperglicemicoДокумент5 страницMetformina 1992 Nao HiperglicemicoCássio SilvaОценок пока нет
- Rice Bran Reduces Weight Gain and Modulates Lipid Metabolism in Rats With High-Energy-Diet-Induced ObesityДокумент10 страницRice Bran Reduces Weight Gain and Modulates Lipid Metabolism in Rats With High-Energy-Diet-Induced ObesityXavier Alvaro Quille MamaniОценок пока нет
- A Physico-Chemical Characterization of A Natural Agglutinin From The Mucus of A Slug Laevicaulis Alte (Ferussac, 1822)Документ12 страницA Physico-Chemical Characterization of A Natural Agglutinin From The Mucus of A Slug Laevicaulis Alte (Ferussac, 1822)TJPRC PublicationsОценок пока нет
- Chlorogenic Acid Improves Obesity and Lipid MetabolismДокумент7 страницChlorogenic Acid Improves Obesity and Lipid MetabolismproluvieslacusОценок пока нет
- Partial or Total Replacement of Fishmeal by Solvent-Extracted Cottonseed Meal in Diets For Juvenile Rainbow Trout (Oncorhynchus Mykiss)Документ8 страницPartial or Total Replacement of Fishmeal by Solvent-Extracted Cottonseed Meal in Diets For Juvenile Rainbow Trout (Oncorhynchus Mykiss)Cherry VelasquezОценок пока нет
- Effect of Alcohol On The Epithelium IntegrityДокумент7 страницEffect of Alcohol On The Epithelium Integrityharis mustofaОценок пока нет
- Dietary and Biliary Phosphatidylcholine Activates PKC in Rat IntestineДокумент12 страницDietary and Biliary Phosphatidylcholine Activates PKC in Rat IntestineTheodora TeddyОценок пока нет
- Jphysiol00649 0467Документ17 страницJphysiol00649 0467Blink SОценок пока нет
- A Comparative Study of Hepatic Mitochondrial Oxygen Consumption in Four Vertebrates by Using Clark-Type ElectrodeДокумент9 страницA Comparative Study of Hepatic Mitochondrial Oxygen Consumption in Four Vertebrates by Using Clark-Type Electrodeerete11Оценок пока нет
- The Arginine Deiminase Pathway in Regulation of Arginine-Ornithine Exchange andДокумент9 страницThe Arginine Deiminase Pathway in Regulation of Arginine-Ornithine Exchange andAvishekh SinhaОценок пока нет
- Effect of Sodium Metabisulphite On Blood Metabolic Status of Wistar RatsДокумент10 страницEffect of Sodium Metabisulphite On Blood Metabolic Status of Wistar Ratsام محمدОценок пока нет
- 1988 - Microencapsulation of Pancreatic Islet Cells - A Bioartificial Endocrine PancreasДокумент6 страниц1988 - Microencapsulation of Pancreatic Islet Cells - A Bioartificial Endocrine PancreasAndrés MenesesОценок пока нет
- Alterations in Myocardial Lipid Metabolism During Lactation in The RatДокумент7 страницAlterations in Myocardial Lipid Metabolism During Lactation in The RatSharkyОценок пока нет
- 5091 PDFДокумент10 страниц5091 PDFNdelooDOnkОценок пока нет
- Lectin Histochemistry For Light and Electron MicrosДокумент12 страницLectin Histochemistry For Light and Electron MicrosInari KikadaОценок пока нет
- Purification and Characterization of An Intracellular 13-Glucosidase From ATCC 393Документ10 страницPurification and Characterization of An Intracellular 13-Glucosidase From ATCC 393Anastasya AmandaОценок пока нет
- CapsaicinДокумент12 страницCapsaicinExlonk Gil PeláezОценок пока нет
- White 1976Документ10 страницWhite 1976poojajain86Оценок пока нет
- Research Article: Increased Yield and Improved Transplantation Outcome of Mouse Islets With Bovine Serum AlbuminДокумент10 страницResearch Article: Increased Yield and Improved Transplantation Outcome of Mouse Islets With Bovine Serum AlbuminCLPHtheoryОценок пока нет
- Mitochondrial Function, Content and ROS Production in Rat Skeletal Muscle: Effect of High-Fat FeedingДокумент7 страницMitochondrial Function, Content and ROS Production in Rat Skeletal Muscle: Effect of High-Fat FeedingRoy WilsonОценок пока нет
- Poonsin2019 PDFДокумент9 страницPoonsin2019 PDFAntonella Acevedo GomezОценок пока нет
- Bioactive Carbohydrates and Dietary FibreДокумент6 страницBioactive Carbohydrates and Dietary FibreJorge MartinezОценок пока нет
- Success Factors for Fish Larval ProductionОт EverandSuccess Factors for Fish Larval ProductionLuis ConceicaoОценок пока нет
- 4ms Scheme For Metallic Materials Part AДокумент31 страница4ms Scheme For Metallic Materials Part APop CatalinОценок пока нет
- Wells Plastics Additives For Polyethylene ExtrusionДокумент4 страницыWells Plastics Additives For Polyethylene ExtrusionmohammedОценок пока нет
- Polymers 14 04269Документ22 страницыPolymers 14 04269Athlur Sai KiranОценок пока нет
- Energy Utilization GuideДокумент29 страницEnergy Utilization GuideYudi SantosoОценок пока нет
- BP 2014 IndexДокумент74 страницыBP 2014 IndexDeepak ShahОценок пока нет
- Strain Improvement of Microorganisms PDFДокумент32 страницыStrain Improvement of Microorganisms PDFsatriomegaОценок пока нет
- Antioxidant Efficacy of Rice Bran Extract On Stabilisation of Ghee Under Accelerated PDFДокумент9 страницAntioxidant Efficacy of Rice Bran Extract On Stabilisation of Ghee Under Accelerated PDFJewel HossenОценок пока нет
- Model 2: General Methods of Assaying Volatile OilsДокумент6 страницModel 2: General Methods of Assaying Volatile OilsAliza EsplanadaОценок пока нет
- Alternative SplicingДокумент25 страницAlternative Splicingtendril123Оценок пока нет
- Alpha-Amylase Assay Procedure (Ceralpha Method) For The Measurement of Plant and Microbial Alpha-AmylasesДокумент20 страницAlpha-Amylase Assay Procedure (Ceralpha Method) For The Measurement of Plant and Microbial Alpha-AmylasesNathanna MateusОценок пока нет
- Tribhuvan Universit1Документ13 страницTribhuvan Universit1PRAJIT BHATTARAIОценок пока нет
- Thermodynamic Modeling Expert Dr. Javeed Ashraf AWANДокумент7 страницThermodynamic Modeling Expert Dr. Javeed Ashraf AWANTalha MughalОценок пока нет
- Nalco Varidos FSK: Cigweld Pty LTDДокумент10 страницNalco Varidos FSK: Cigweld Pty LTDPouria KazemzadehОценок пока нет
- Towards Paired and Coupled Electrode Reactions For Clean Organic Microreactor ElectrosynthesisДокумент18 страницTowards Paired and Coupled Electrode Reactions For Clean Organic Microreactor ElectrosynthesisjorgchanОценок пока нет
- LANXESS FF Aroma Ingredients - Update 2023-10SДокумент2 страницыLANXESS FF Aroma Ingredients - Update 2023-10ScОценок пока нет
- To-GE KC-7 Methyl Salicylate 15% Menthol 7% Hydro Alcoholic Clear GelДокумент1 страницаTo-GE KC-7 Methyl Salicylate 15% Menthol 7% Hydro Alcoholic Clear GelAnand Rao100% (1)
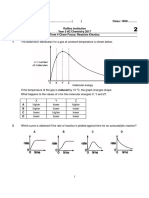
- 2707 Chemistry Paper With Solutions MorningДокумент7 страниц2707 Chemistry Paper With Solutions MorningSubham roushanОценок пока нет
- Haloalkanes and HaloarenesДокумент9 страницHaloalkanes and HaloarenesManish kumarОценок пока нет
- D2qo00551d 3598..3623 ++Документ26 страницD2qo00551d 3598..3623 ++Shaswata KunduОценок пока нет
- Chemistry of Cocoa Beans and ProductsДокумент70 страницChemistry of Cocoa Beans and ProductsdindaОценок пока нет
- Q1.The Diagram Shows A Mitochondrion.: Feversham CollegeДокумент56 страницQ1.The Diagram Shows A Mitochondrion.: Feversham CollegeZaksd100% (1)
- 2017 Y5 T4 Chem Focus - KineticsДокумент4 страницы2017 Y5 T4 Chem Focus - KineticsxmxmxmxmxmОценок пока нет
- Effect of potassium bisulphite as a food preservativeДокумент7 страницEffect of potassium bisulphite as a food preservativesriramОценок пока нет
- Vetiver (Type of Grass)Документ12 страницVetiver (Type of Grass)p6a4nduОценок пока нет
- Printed Thinking Map From 5Документ16 страницPrinted Thinking Map From 5Mohd Rizal LakimОценок пока нет
- Lesson 1 Introduction To Organic Chemistry PDFДокумент4 страницыLesson 1 Introduction To Organic Chemistry PDFdela2Оценок пока нет
- The Sulfur Cycle: by Guest On 09 September 2021Документ45 страницThe Sulfur Cycle: by Guest On 09 September 2021Miguel MoreiraОценок пока нет
- Worksheet On Act#3 Alcoholic FermentationДокумент4 страницыWorksheet On Act#3 Alcoholic FermentationKrizia CabayacruzОценок пока нет
- Effective Microorganisms (Em Technology and Bokashi)Документ177 страницEffective Microorganisms (Em Technology and Bokashi)Rommel SacramentoОценок пока нет
- Determination of Sugars in Molasses by HPLCДокумент12 страницDetermination of Sugars in Molasses by HPLCNatzi MonsalvoОценок пока нет