Вам также может понравиться
- Tmpa381 TMPДокумент22 страницыTmpa381 TMPFrontiersОценок пока нет
- EJPB - Volume 24 - Issue 4 - Pages 889-900Документ12 страницEJPB - Volume 24 - Issue 4 - Pages 889-900Mourad TaibiОценок пока нет
- Phylogeny and Evolution of Basils and Allies (Ocimeae, Labiatae) Based On Three Plastid DNA RegionsДокумент23 страницыPhylogeny and Evolution of Basils and Allies (Ocimeae, Labiatae) Based On Three Plastid DNA RegionslinubinoiОценок пока нет
- A.C. Zeven J. Waninge Th. Van Hintum S.P. Singh - Phenotypic Variation in A Core Collection of Common Bean (Phaseolus Vulgaris L.) in The NetherlaДокумент14 страницA.C. Zeven J. Waninge Th. Van Hintum S.P. Singh - Phenotypic Variation in A Core Collection of Common Bean (Phaseolus Vulgaris L.) in The NetherlaNicholas ViniciusОценок пока нет
- tmpAC TMPДокумент12 страницtmpAC TMPFrontiersОценок пока нет
- Quantitative Genetics of RiceДокумент7 страницQuantitative Genetics of RiceSenguttuvel PonnuvelОценок пока нет
- Regeneration and Transformation of An Elite Inbred Line of Maize (Zea Mays L.), With A Gene From Bacillus ThuringiensisДокумент7 страницRegeneration and Transformation of An Elite Inbred Line of Maize (Zea Mays L.), With A Gene From Bacillus ThuringiensisCitlalli CedilloОценок пока нет
- Genetic Studies in Upland Cotton.Документ10 страницGenetic Studies in Upland Cotton.Barkat2012Оценок пока нет
- Interspecific Hybridization in Gossypium L.: Characterization of Progenies With Different Ploidy-Confirmed Multigenomic BackgroundsДокумент7 страницInterspecific Hybridization in Gossypium L.: Characterization of Progenies With Different Ploidy-Confirmed Multigenomic BackgroundsnarayananОценок пока нет
- Rahulcreditseminar 161110121841Документ70 страницRahulcreditseminar 161110121841narayananОценок пока нет
- PNAS 1997 Yu 9226 31Документ6 страницPNAS 1997 Yu 9226 31Dani Pazmiño BonillaОценок пока нет
- Dissection of The Genetic Basis of Heterosis in An Elite Maize Hybrid by QTL Mapping in An Immortalized F2 PopulationДокумент8 страницDissection of The Genetic Basis of Heterosis in An Elite Maize Hybrid by QTL Mapping in An Immortalized F2 PopulationvahidvasharОценок пока нет
- TMP 9 E37Документ8 страницTMP 9 E37FrontiersОценок пока нет
- Ijasr - Diallel Cross Analysis For Earliness, Yield, Its Components andДокумент12 страницIjasr - Diallel Cross Analysis For Earliness, Yield, Its Components andTJPRC PublicationsОценок пока нет
- Weider Stability CMS 20091Документ9 страницWeider Stability CMS 20091yustianaОценок пока нет
- Transferring Hessian Fly Resistance from Aegilops geniculata to Durum WheatДокумент6 страницTransferring Hessian Fly Resistance from Aegilops geniculata to Durum WheatMuhammad Luqman KuswardanaОценок пока нет
- BCAT-based Marker For Marker-Assisted Selection in Vietnam Cucumber BreedingДокумент5 страницBCAT-based Marker For Marker-Assisted Selection in Vietnam Cucumber BreedingShailendra RajanОценок пока нет
- Helia - 2018 - 0013 - Stability Analysis of Noval Cms Sources - BlindДокумент44 страницыHelia - 2018 - 0013 - Stability Analysis of Noval Cms Sources - BlindMarija Kraljevic-BalalicОценок пока нет
- The Production of Callus Capable of Plant Regeneration From Immature Embryos Zea MaysДокумент11 страницThe Production of Callus Capable of Plant Regeneration From Immature Embryos Zea Mayssiti.dede1973Оценок пока нет
- Gene effects and sequential models for sesame yield traitsДокумент12 страницGene effects and sequential models for sesame yield traitsJawahar JawaharОценок пока нет
- AndyGWAS-RevisedManuscript FileДокумент26 страницAndyGWAS-RevisedManuscript FileZeleke WondimuОценок пока нет
- Robles 2005 ABC ModelДокумент11 страницRobles 2005 ABC ModelJegan Passiflora EdulisОценок пока нет
- Genetic Stability Among Date Palm Plantlets Regene PDFДокумент9 страницGenetic Stability Among Date Palm Plantlets Regene PDFngoclinhtu9816Оценок пока нет
- NY Regents Living Environment Exam GuideДокумент28 страницNY Regents Living Environment Exam Guidemeg carrОценок пока нет
- Molecular Markers For Sex Determination in Papaya (Carica Papaya L.)Документ5 страницMolecular Markers For Sex Determination in Papaya (Carica Papaya L.)FrancoОценок пока нет
- Rinaudo 2004Документ11 страницRinaudo 2004Ruyu WangОценок пока нет
- Inheritance of Fruit Flesh Colour in Some Botanical Varieties of Muskmelon, Cucumis MeloДокумент5 страницInheritance of Fruit Flesh Colour in Some Botanical Varieties of Muskmelon, Cucumis MeloShailendra RajanОценок пока нет
- Karakteristik Bunga TelangДокумент7 страницKarakteristik Bunga TelangRahmahPuspitaSariОценок пока нет
- Characterization of Myostatin Gene and Identification of Snps For Diversity AnalysisДокумент9 страницCharacterization of Myostatin Gene and Identification of Snps For Diversity AnalysisnikhilsathwikОценок пока нет
- 1Документ34 страницы1ahmredОценок пока нет
- Genetic Characterization of PiliДокумент11 страницGenetic Characterization of PiliEunice ChavezОценок пока нет
- Genetics and Cytogenetics of The PotatoДокумент29 страницGenetics and Cytogenetics of The PotatoJOSE FRANCISCO FRANCO NAVIAОценок пока нет
- Heterosis For The Improvement of Oil QualityДокумент6 страницHeterosis For The Improvement of Oil QualityShujaul Mulk KhanОценок пока нет
- Expression of Anthocyanins and Proanthocyanidins After Transformation of Alfalfa With Maize LCДокумент16 страницExpression of Anthocyanins and Proanthocyanidins After Transformation of Alfalfa With Maize LC10sgОценок пока нет
- Generation Mean Analysis of Some Agronomic Traits in HB42xSabini Barley CrossДокумент9 страницGeneration Mean Analysis of Some Agronomic Traits in HB42xSabini Barley CrossPremier PublishersОценок пока нет
- Cáceres 2009Документ14 страницCáceres 2009Manu Gustani BussОценок пока нет
- Aphyosemion and Fundulopanchax 1999Документ10 страницAphyosemion and Fundulopanchax 1999Mario Manuel Rueda ViguerasОценок пока нет
- Estimates of Genetic and Environmental Variability in SoybeansДокумент1 страницаEstimates of Genetic and Environmental Variability in SoybeansFitria DelaОценок пока нет
- Plant GeneticsДокумент9 страницPlant GeneticsKauntvladОценок пока нет
- SVUIJAS - Volume 4 - Issue 4 - Pages 79-89Документ11 страницSVUIJAS - Volume 4 - Issue 4 - Pages 79-89Safaa IsmailОценок пока нет
- Phylogeny of Stemphylium SPP Based On ITS and Glyceraldehyde 3 Phosphate Dehydrogenase Gene SequencesДокумент14 страницPhylogeny of Stemphylium SPP Based On ITS and Glyceraldehyde 3 Phosphate Dehydrogenase Gene SequencesJelle van CampenhoutОценок пока нет
- Sex DeterminationДокумент11 страницSex DeterminationJulio SubagioОценок пока нет
- Somatic Embryogenesis and Plant RegenerationДокумент6 страницSomatic Embryogenesis and Plant RegenerationmacroendrixОценок пока нет
- Allium CepaДокумент9 страницAllium CepaNuke Nur HidayatiОценок пока нет
- Artigo Reference Genes ArabidopsisДокумент13 страницArtigo Reference Genes ArabidopsisAline CunhaОценок пока нет
- Write Up GP 591Документ21 страницаWrite Up GP 591rickos90Оценок пока нет
- Micropropagation of Medicinal PlantsДокумент23 страницыMicropropagation of Medicinal Plants존 셔윈Оценок пока нет
- Identification of Maintainer and Restorer Lines in LOCAL AROMATIC RICE (Oryza Sativa)Документ12 страницIdentification of Maintainer and Restorer Lines in LOCAL AROMATIC RICE (Oryza Sativa)Rushabh MeherОценок пока нет
- Biological Control: Liliana Hoyos-Carvajal, Sergio Orduz, John BissettДокумент8 страницBiological Control: Liliana Hoyos-Carvajal, Sergio Orduz, John BissettAde PutОценок пока нет
- PGR Arun 2011Документ4 страницыPGR Arun 2011arunprabhu_dhanapalОценок пока нет
- Agri Sci - Jasr - Assessment of Herbicide AlachlorДокумент8 страницAgri Sci - Jasr - Assessment of Herbicide AlachlorTJPRC PublicationsОценок пока нет
- Jee96 0914Документ11 страницJee96 0914Aimer GuzmanОценок пока нет
- Soneratia AlbaДокумент5 страницSoneratia AlbaRafil FriasmarОценок пока нет
- Inbred Lines Maize and Their Molecular MarkersДокумент10 страницInbred Lines Maize and Their Molecular Markersjaber AzimОценок пока нет
- Euphyticapaper PDFДокумент8 страницEuphyticapaper PDFAulia FebriantiОценок пока нет
- Elsevier PollinationДокумент12 страницElsevier PollinationChris MaliachovasОценок пока нет
- Development of A Groundnut Core Collection Using Taxonomical, Geographical and Morphological DescriptorsДокумент10 страницDevelopment of A Groundnut Core Collection Using Taxonomical, Geographical and Morphological DescriptorsJonahОценок пока нет
- Unbeatable Strategy Constraint and Coevo PDFДокумент9 страницUnbeatable Strategy Constraint and Coevo PDFmunazzaОценок пока нет
- tmpAE4A TMPДокумент12 страницtmpAE4A TMPFrontiersОценок пока нет
- Analysis of Genetic Diversity in Germpla PDFДокумент4 страницыAnalysis of Genetic Diversity in Germpla PDFmangeshdudhe2458Оценок пока нет
- 10 1 1 177 7561Документ5 страниц10 1 1 177 7561mangeshdudhe2458Оценок пока нет
- SABRAO J Breed Genet 49-1-83 93 DudheДокумент11 страницSABRAO J Breed Genet 49-1-83 93 Dudhemangeshdudhe2458Оценок пока нет
- Sunflower Breeding Strategy For Resistance To Downy Mildew Disease in IndiaДокумент6 страницSunflower Breeding Strategy For Resistance To Downy Mildew Disease in Indiamangeshdudhe2458Оценок пока нет
- Importance of Meta-Qtl Analysis For Crop ImprovementДокумент5 страницImportance of Meta-Qtl Analysis For Crop Improvementmangeshdudhe2458Оценок пока нет
- In Vitro Screening of Wheat Genotypes For Drought Tolerance On The Basis of Callus InductionДокумент5 страницIn Vitro Screening of Wheat Genotypes For Drought Tolerance On The Basis of Callus Inductionmangeshdudhe2458Оценок пока нет
- Analysis of Genetic Diversity in Germplasm Accessions of Sunflower (Helianthus Annuus L.)Документ4 страницыAnalysis of Genetic Diversity in Germplasm Accessions of Sunflower (Helianthus Annuus L.)mangeshdudhe2458Оценок пока нет
- Study of Gene Action For Restorer Lines InsunflowerДокумент6 страницStudy of Gene Action For Restorer Lines Insunflowermangeshdudhe2458Оценок пока нет
- Dr. Mangesh Y. Dudhes Published PaperДокумент8 страницDr. Mangesh Y. Dudhes Published Papermangeshdudhe2458Оценок пока нет
- Dr. Mangesh Y. Dudhes PosterДокумент1 страницаDr. Mangesh Y. Dudhes Postermangeshdudhe2458Оценок пока нет
- Dr. Mangesh Y. Dudhes PaperДокумент3 страницыDr. Mangesh Y. Dudhes Papermangeshdudhe2458Оценок пока нет
- List of Indian Plants by Name and Botanical NameДокумент3 страницыList of Indian Plants by Name and Botanical NameNAYAN JYOTI KALITAОценок пока нет
- 1P13 - Innaka Ageng 378-384Документ7 страниц1P13 - Innaka Ageng 378-384Dwiky Riza MhendraОценок пока нет
- Tubang BakodДокумент4 страницыTubang BakodDei HernandezОценок пока нет
- TBB's Wedding Color Guide + ThemesДокумент33 страницыTBB's Wedding Color Guide + ThemesKathleenJoyEnSagun100% (1)
- Elodea Lab PDFДокумент5 страницElodea Lab PDFVikash kumarОценок пока нет
- Chapter 1 - Chemical Process Diagrams: Department of Chemical Engineering West Virginia UniversityДокумент38 страницChapter 1 - Chemical Process Diagrams: Department of Chemical Engineering West Virginia UniversitycatherineОценок пока нет
- Subhayu Ghosh - MOCK TEST 2Документ31 страницаSubhayu Ghosh - MOCK TEST 2Subhayu GhoshОценок пока нет
- Owb FactsheetДокумент17 страницOwb FactsheetRahmania OktaОценок пока нет
- O Level 5090 Scheme of WorkДокумент50 страницO Level 5090 Scheme of WorkBalakrishnan Marappan100% (2)
- Anthurium Micropropagation From Shoot Tip Explants PDFДокумент5 страницAnthurium Micropropagation From Shoot Tip Explants PDFBrij Mohan SinghОценок пока нет
- Crisp Cookbook Recipes for Whirlpool Microwave OvenДокумент8 страницCrisp Cookbook Recipes for Whirlpool Microwave OvenJudit Maria BerzeОценок пока нет
- Home Composting Brochure Cwmi - Css.cornell - EduДокумент4 страницыHome Composting Brochure Cwmi - Css.cornell - EduPennsylvania Association for Sustainable AgricultureОценок пока нет
- Insecticidal Property of Extract of Piper NigrumДокумент6 страницInsecticidal Property of Extract of Piper NigrumOliver TalipОценок пока нет
- A321 Technical Training Manual CFM56-5B Engine FamiliarizationДокумент92 страницыA321 Technical Training Manual CFM56-5B Engine FamiliarizationAnas AbandehОценок пока нет
- ELS Group Work - Plant and Animal ReproductionДокумент1 страницаELS Group Work - Plant and Animal ReproductionMichael Paul SinsuanОценок пока нет
- Botrytis Bunch Rot of Grape (Botrytis Cinerea) - Pest Management - B.C. Ministry of AgricultureДокумент4 страницыBotrytis Bunch Rot of Grape (Botrytis Cinerea) - Pest Management - B.C. Ministry of AgricultureSiva KumarОценок пока нет
- Survivalist CookbookДокумент25 страницSurvivalist CookbookJohn H McDonald100% (1)
- Royal Botanic GardensДокумент18 страницRoyal Botanic GardensRAISAОценок пока нет
- Study On Physico-Chemical Properties of Developed Ready To Use Health Beverage Containing CoffeeДокумент4 страницыStudy On Physico-Chemical Properties of Developed Ready To Use Health Beverage Containing CoffeeAnonymous izrFWiQОценок пока нет
- Etta S Lunch MenuДокумент3 страницыEtta S Lunch MenueatlocalmenusОценок пока нет
- Meeting 14 Toefl Simulation 2 Reading PDFДокумент10 страницMeeting 14 Toefl Simulation 2 Reading PDFSalsabill NahdahОценок пока нет
- CAMAG Application Notes HerbalsДокумент2 страницыCAMAG Application Notes HerbalsGustavo RuizОценок пока нет
- Enfagrow A+ and Sustagen product inventory and returnsДокумент4 страницыEnfagrow A+ and Sustagen product inventory and returnsEmeng CatShopОценок пока нет
- Crome969 HeartwoodДокумент8 страницCrome969 HeartwoodAlcatrazz Albion OnlineОценок пока нет
- Pyrography Pen Tips: Lora S. IrishДокумент38 страницPyrography Pen Tips: Lora S. Irishalbertanton99100% (1)
- Landscape Architecture GlossaryДокумент10 страницLandscape Architecture GlossaryCar TejadaОценок пока нет
- Herbs Magical Properties For Protection, Healing and PovertyДокумент18 страницHerbs Magical Properties For Protection, Healing and PovertyTom ChordОценок пока нет
- A. Multiple Choice Questions. Choose The Best Answer!Документ3 страницыA. Multiple Choice Questions. Choose The Best Answer!Manafe LexОценок пока нет
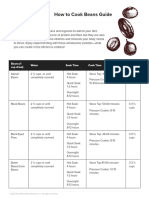
- How To Cook Beans GuideДокумент6 страницHow To Cook Beans GuideIvan Kovač KrčelićОценок пока нет