Академический Документы
Профессиональный Документы
Культура Документы
Celula Vegetal
Загружено:
Fernando MéndezОригинальное название
Авторское право
Доступные форматы
Поделиться этим документом
Поделиться или встроить документ
Этот документ был вам полезен?
Это неприемлемый материал?
Пожаловаться на этот документАвторское право:
Доступные форматы
Celula Vegetal
Загружено:
Fernando MéndezАвторское право:
Доступные форматы
La teora celular actualmente se puede resumir de la siguiente forma : 1.
Todos los organismos vivos estn formados por clulas y productos celulares. 2. Slo se forman clulas nuevas a partir de clulas preexistentes. 3. La informacin gentica que se necesita durante la vida de las clulas y la que se requiere para la produccin de nuevas clulas se transmite de una generacin a la siguiente. 4. Las reacciones qumicas de un organismo, esto es su metabolismo, tienen lugar en las clulas.
CLULAS VEGETALES Y ANIMALES
Tanto las clulas de las plantas como las de los animales son eucariticas, sin embargo presentan algunas diferencias:
1. Las clulas vegetales presentan una pared celular celulsica, rgida que evita
cambios de forma y posicin. 2. Las clulas vegetales contienen plastidios, estructuras rodeadas por una membrana, que sintetizan y almacenan alimentos. Los ms comunes son los cloroplastos. 3. Casi todas las clulas vegetales poseen vacuolas, que tienen la funcin de transportar y almacenar nutrientes, agua y productos de desecho. 4. Las clulas vegetales complejas, carecen de ciertos organelos, como los centriolos y los lisosomas. Las plantas son organismos multicelulares formados por millones de clulas con funciones especializadas. Sin embargo, todas las clulas vegetales poseen una organizacin comn: tienen un ncleo, un citoplasma y organelos subcelulares; los cuales se encuentran rodeados por una membrana que establece sus lmites. As como una pared celular que rodea el protoplasto (ncleo + citoplasma con sus inclusiones).
LA PARED CLULAR
Aunque las clulas vegetales y animales son muy parecidas, las clulas vegetales tienen una pared rgida de celulosa, que le brinda proteccin, sin impedir la difusin de agua y iones desde el medio ambiente hacia la membrana plasmtica, que es la verdadera barrera de permeabilidad de la clula. Una pared celular primaria tpica, de una dicotilednea est formada por 25-30 % de celulosa, 15-25 % de hemicelulosa, 35 % de pectina y 5-10 % de protenas (extensinas y lectinas), en base al peso seco. La constitucin molecular y estructural precisa de la pared celular, depende del tipo de clula, tejido y especie vegetal. La pared primaria es delgada ( de 1 a 3 micras de grosor) y se forma cuando la clula crece, ejemplo de esta la tenemos en clulas jvenes en crecimiento, en el tejido parenquimtico, en el clornquima, epidermis, etc. La membrana celular est fuertemente adherida a la pared celular, debido a la presin de turgencia provocada por los fluidos intracelulares. Literalmente podemos decir que las clulas se encuentran abombadas, empujndose entre ellas; en otras palabras se encuentran infladas por una presin hidrosttica.
Las macromolculas de celulosa, en la pared celular esta formada por unidades de glucosa (un azcar de 6 carbonos) enlazadas covalentemente, formando una estructura en forma de cinta aplanada, que puede tener de 0,25 a 5 micras de largo. Entre 40 a 70 de estas cadenas se mantienen unidas mediante enlaces de hidrgeno, entre los grupos OH de los residuos de glucosa, formando una estructura cristalina llamada microfibrilla, que tiene aproximadamente 3 nm de dimetro. La celulosa es muy estable qumicamente e insoluble. Las microfibrillas tienen una alta fuerza tensional, que actua reforzando la pared. Grupos de microfibrillas se disponen como los alambres en un cable, formando macrofibrillas. Las macrofibrillas son los componentes ms importantes de la pared celular y se mantienen unidas mediante otros componentes de la pared celular, como son las macromolculas de hemicelulosa y pctina. Estas sustancias pegan toda la estructura, en capas de fibras. Las primeras microfibrillas que se depositan en la pared celular, forman una red con disposicin transversal. Pero, cuando la presin de turgencia produce la extensin celular y la pared crece en rea superficial, la otra capa de microfibrillas se deposita paralelamente, al eje longuitudinal de la clula. El efecto final es una apariencia entramada de varias capas. Dos clulas adyacentes se mantienen unidas mediante la lmina media, la que se encuentra formada principalmente por sustancias pecticas, que cementan las paredes primarias, a ambos lados de la lmina media. Nosotros podemos extraer la pectina de frutos verdes, como por Ej. el mango y hacer jalea. En muchas plantas posteriormente se puede depositar una pared celular secundaria, que imparte rigidez y fortaleza al tejido, s se deposita lignina. Por ejemplo los troncos de los rboles, tienen clulas con gruesas paredes celulares secundarias. Las plantas multicelulares, se conectan a travs de pequeas perforaciones que comunican las clulas adyacentes, denominadas campos de punteaduras primarias, a travs de los cuales pasan cordones citoplasmticos denominados plasmodesmos. A pesar de que son muy pequeos para que lo atraviesen organelos celulares, sin embargo las conexiones citoplasmticas permiten la transferencia de sustancias de una clula a otra. La membrana plasmtica es continua y se extiende de una clula a la otra a travs de los plasmodesmos, constituyendo lo que se conoce como simplasto ; mientras que el conjunto de las paredes celulares de un tejido, ms los espacios intercelulares, se denomina apoplasto. La pared celular es muy permeable a diferentes sustancias, permitiendo el paso de agua y solutos; aunque la verdadera barrera que controla la permeabilidad, al igual que en las clulas animales, es la membrana plasmtica o plasmalema
LA MEMBRANA PLASMTICA
La membrana plasmtica, tanto de las clulas procariticas como eucariticas, son bsicamente similares. En ambos casos, regula el flujo de sustancias disueltas hacia adentro y hacia afuera de la clula. La smosis, que funciona debido a que el agua pasa a travs de las membranas ms rpido que los solutos, regula el flujo de agua. Las membranas plasmticas tienen aproximadamente 50% de fosfolpidos y 50% de protenas. La estructura en tres capas de las membranas celulares, consiste de una doble capa de fosfolpidos, con los grupos hidrofbos(no afines al agua) mirando hacia el centro y los grupos hidroflicos (afines al agua) orientados hacia las partes externas de la bicapa lpidica. Las molculas de protenas, flotan en la bicapa lipdica, con sus terminaciones hidroflicas penetrando en ambas superficies de la membrana, lo que se conoce como el modelo de mosaico fluido, propuesto por Singer y Nicolson (1972). Se sabe que en las membranas existen dos tipos de protenas : las protenas integrales (intrnsecas) y las protenas perifricas (extrnsecas).
Cuando se estudia la membrana plasmtica mediante el microscopio electrnico, despus de haber sido apropiadamente fijada con tetrxido de osmio, las capas de protenas se observan como dos lneas densas (oscuras), con un espacio claro entre ellas. Las lneas oscuras tienen un espesor de aproximadamente 2,5 a 3,5 nm y la lnea clara tiene aproximadamente 3,5 nm, para un grosor de aproximadamente 10 nm o100 . La que se conoce como la unidad de membrana. Esto no significa que todas las membranas sean iguales; ya que ellas pueden presentar diferentes caractersticas de permeabilidad. El hecho de que una sustancia pueda atravesar la membrana de un cloroplasto, no significa que lo pueda hacer tambin a travs de una membrana mitocondrial. Las membranas poseen la propiedad de ser selectivas, lo que indica que cada tipo de membrana tiene caractersticas moleculares particulares, que les permite funcionar bajo sus propias condiciones. Todas las membranas biolgicas que rodean las clulas, ncleos, vacuolas, mitocondrias, cloroplastos y otros organelos celulares son selectivamente permeables. Las membranas son muy permeables a las molculas de agua y ciertos gases, incluyendo el oxgeno y el dixido de carbono ; mientras que otras molculas pueden tener problemas para atravesar las membranas, debido a su tamao, polaridad y solubilidad en lpidos. Los iones y las molculas polares (con carga elctrica), tienden a moverse a travs de la parte proteica de la membrana. Muchas sustancias se mueven mediante difusin simple, por un proceso de transporte pasivo, de zonas de mayor a menor concentracin. Sin embargo, en los seres biticos muchas sustancias atraviesan la membrana mediante transporte activo, movindose en contra de un gradiente de concentracin, y con la utilizacin de energa metablica por la clula , en forma de ATP (adenosin trifosfato), el cual es aportado por la respiracin
EL PROTOPLASTO
El contenido del protoplasto, se puede dividir en tres partes fundamentales: citoplasma, ncleo y vacuola(s); as mismo se encuentran sustancias ergsticas y rganos de locomocin. Todas las clulas eucariticas, al menos cuando jvenes poseen un ncleo ; el cual puede desaparecer en los tubos cribosos y en otras clulas vegetales, en la medida que maduran. El protoplasto se encuentra ausente en los elementos xilemticos maduros ( vasos y traqueidas). La presencia de vacuolas y sustancias ergsticas, es una caracterstica de las clulas de hongos y de las plantas.
El citoplasma (plasma fundamental), tiene una consistencia viscosa y est compuesto de una mezcla heterognea de protenas (enzimas) y es el lugar donde ocurren importantes reacciones metablicas, como la gluclisis. Debido a su naturaleza coloidal, el citoplasma sufre cambios de estado, puede pasar de sol (fluido) a gel (parecido a la gelatina). El citosol, es la matriz fluida en la que los organelos se encuentran suspendidos, est organizado en una red tridimensional de protenas fibrosas, llamadas citoesqueleto. El citoesqueleto es mucho ms organizado, que la sopa clara que nos podemos imaginar. Los elementos del citoesqueleto son: los microtbulos y los microfilamentos. Los microtbulos son filamentos cilndricos, huecos que tienen un dimetro externo de 25 nm y varias micras de longitud. Las paredes de los microtbulos, estn formadas por filamentos proticos lineares o en espiral de aproximadamente 5 nm de dimetro y estos estn compuestos de 13 subunidades. En el centro de un microtbulo se encuentra un lumen (rea vaca); sin embargo se pueden observar bastones o puntos. Los microtbulos estan compuestos por molculas esfricas de una protena llamada tubulina. Los microtbulos pueden formarse o descomponerse rpidamente a conveniencia, y se encuentran formando parte de estructuras celulares que facilitan el movimiento, como el huso mittico y los flagelos. La colquicina, un alcaloide del clquico (Colchicum autumnale), destruye la organizacin de los microtbulos, impidiendo la formacin del huso acromtico durante la mitosis celular. Por lo que la colquicina se ha utilizado en gentica, en la obtencin de clulas poliploides. Los microfilamentos son estructuras ms pequeas, pero slidas de 5 a 7 nm de dimetro, que actan solos o conjuntamente con los microtbulos para producir movimiento celular. Estos tambin estn formados por protenas, especficamente la protena actina, la que con la miosina son tambin constituyentes del tejido muscular de los animales. Los microfilamentos causan el movimiento de corriente citoplasmtica o ciclosis, la que ocurre en muchas clulas vegetales, como en las algas Chara y Nitella, donde se han reportado velocidades de 75 por segundo. En las hojas de la Elodea canadensis, se observa muy bien la ciclosis, que produce un movimiento de los organelos celulares, de una forma helicoidal, de un lado hacia abajo y del otro lado hacia arriba. Los microfilamentos tambin juegan un papel importante en el crecimiento del tubo polnico y en el movimiento ameboidal. En el citoplasma se encuentra un sistema de endomembranas, que incluye al retculo endoplasmtico, el aparato de Golgi, la envoltura nuclear y otros organelos celulares y membranas (tales como los microcuerpos, esferosomas y membrana vacuolar), que tienen sus orgenes en el retculo endoplasmtico o en el aparato de Golgi. La membrana celular que ya la hemos estudiado, se considera como una entidad separada; aunque su crecimiento se debe a la adicin de vesculas por el aparato de Golgi. Las mitocondrias y plastidios se encuentran rodeados por una doble membrana, que se parece al sistema de endomembranas; aunque estos organelos se autoduplican, por lo que no estn relacionados al sistema de endomembranas. As mismo, los ribosomas, los microtbulos y los microfilamentos, no forman parte del sistema de endomembranas. El retculo endoplasmtico (RE o ER, del ingls endoplasmic reticulum) es un sistema multirramificado de sacos membranosos planos, denominados cisternas, que presentan la tpica estructura de unidad de membrana. El RE es continuo con la membrana externa de la envoltura nuclear, a la que se une en las cercanas del ncleo. El RE puede tener ribosomas, que se encuentran unidos como lo hacen los botones a un pedazo de tela, y se conoce como RE rugoso o puede carecer de ribosomas y se llama RE liso. El RE rugoso sintetiza lpidos de membrana y protenas de secrecin; mientras que el RE liso est implicado tambin en la produccin de lpidos y en la modificacin y transporte de las protenas sintetizadas
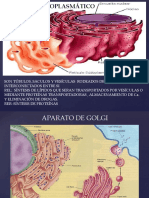
en el RE rugoso. Los ribosomas, observados en una micrografa electrnica a bajo aumento, aparecen como puntos negros, redondos sobre el RE, pero a altos aumentos se observa que estn formados por un cuerpo pequeo esfrico y un cuerpo cncavo grande, tienen de 20 a 30 nm de grosor. Frecuentemente aparecen formando agregados caractersticos que reciben el nombre de polisomas. Los ribosomas son partculas de ribonucleoprotenas (contienen protenas y cido ribonucleco), donde se produce la sntesis de protenas a partir de aminocidos, mediante el mecanismo de la traduccin, de la informacin gentica contenida en el cido ribonucleico mensajero (ARNm). En una clula pueden existir miles de ribosomas, con una capacidad de sntesis prodigiosa, ya que cada ribosoma puede producir una molcula de protena por minuto. Complejos de Golgi o aparato de Golgi esta relacionado con el RE; ste sistema de membranas est compuesto por conjuntos de sacos de Golgi, aplanados y llenos de fluido. Se observan como membranas aplanadas, parecidas a una pila de cachapas. En los extremos de estas membranas aplanadas o cisternas, se pueden observar vesculas que contienen las macromolculas que se usan para la construccin de las membranas y la pared celular. Tanto los polisacridos hemicelulosa y pectina, como la protena de la pared celular (extensina) son sintetizados y procesados en el interior de las vesculas de secrecin del aparato de Golgi o dictiosoma. Cada aparato de Golgi tiene 4 a 6 cisternas con una separacin de 10 nm; no obstante algunas algas pueden tener de 20 a 30. El aparato de Golgi puede tener otras funciones adems de contribuir al crecimiento del plasmalema y transporte de material a la pared celular, como es la de segregar muclago en la parte externa de la punta de la raz, que acta como un lubricante permitiendo su movimiento entre las partculas del suelo. El aparato de Golgi es abundante en muchas clulas secretoras. Los dictiosomas no son estructuras permanentes y en caso de necesidad se forman de novo por el retculo endoplasmtico. Microcuerpos, peroxisomas, glioxisomas. Los microcuerpos son organelos esfricos, rodeados por una sola unidad de membrana. Su dimetro vara de 0,5 a 1,5 y tienen un interior granular; algunas veces con inclusiones cristalinas de protenas. Se originan a partir del RE, formando parte del sistema de endomembranas. Los peroxisomas son organelos esfricos, especializados en reacciones de oxidacin. La enzima catalasa, constituye casi el 40% de las protenas totales del peroxisoma, esta enzima descompone el perxido de hidrgeno en agua y oxgeno. En las plantas se conocen los peroxisomas foliares, como organelos de la foto respiracin. Los glioxisomas se encuentran en semillas de oleaginosas, y contienen las enzimas que ayudan a convertir las grasas almacenadas, en carbohidratos que son translocados a la planta joven para su crecimiento. Los glioxisomas contienen las enzimas del ciclo del cido gliclico. Plastidios. Adems del ncleo y las vacuolas, los plastidios constituyen los organelos ms conspicuos de una clula vegetal. Los plastidios estn rodeados por una doble membrana, con una estructura interna constituida por un sistema de membranas, separadas por una matriz de naturaleza proteica llamada estroma. Los plastidios tienen ADN (DNA) con una estructura similar al encontrado en clulas procariticas, as como ribosomas, embebidos en el estroma. Todos los plastidios se desarrollan a partir de proplastidios, que son cuerpos pequeos encontrados en plantas que crecen tanto en la luz como en la oscuridad. Se dividen por fisin o biparticin, como lo hacen las mitocondrias y los organismos procarites. Los plastidios incoloros se conocen como leucoplastos, contienen enzimas responsables de la sntesis del almidn. Los leucoplastos mejor conocidos son los amiloplastos, que almacenan granos de almidn, como los encontrados en la raz de la yuca, el tubrculo de la papa, en granos de cereales, etc. Otros leucoplastos pueden almacenar protenas, se conocen como proteinoplastos. Los cromoplastos son
organelos coloreados, especializados en sintetizar y almacenar pigmentos carotenoides (rojo, anaranjado y amarillo), estos son el origen de los colores de muchos frutos, flores y hojas, por ej. la piel del tomate, la raz de zanahoria, etc. Los cromoplastos se originan a partir de cloroplastos jvenes o de cloroplastos maduros, por divisin. Los cloroplastos son plastidios que contienen los pigmentos verdes clorofila a y b, as como carotenoides de color anaranjado y xantofilas amarillas, son caractersticos de los seres fotoauttrofos, que poseen la maquinaria enzimtica para transformar la energa solar en energa qumica, a travs de la fotosntesis. Los cloroplastos son caractersticos de las clulas del mesfilo foliar, poseen una doble membrana que los asemeja a las mitocondrias. Tienen una membrana externa y otra interna, el espacio delimitado por la membrana interna est ocupado por un material amorfo, parecido a un gel, rico en enzimas, denominado estroma. Contiene las enzimas que realizan la fijacin o reduccin del CO2, convirtindolo en carbohidratos, como el almidn. La membrana interna de los cloroplastos tambin engloba un tercer sistema de membranas, que consta de sacos planos llamados tilacoides, en los cuales la energa luminosa se utiliza para oxidar el agua y formar ATP (compuesto rico en energa) y NADPH (poder reductor), usados en el estroma para convertir el CO2 en carbohidratos. En ciertas partes de los cloroplastos, los tilacoides se disponen como monedas apiladas, denominados grana, pero en el estroma permanecen aislados. Los cloroplastos tienen forma elptica, con un dimetro de 5 a 10 y su nmero puede variar de 20 a 100 por clula vegetal. Durante la ciclosis se mueven libremente en el citoplasma. Ellos responden directamente a la energa solar, para llevar a cabo la fotosntesis, orientndose perpendicularmente a los rayos de luz; sin embargo s la energa lumnica es muy fuerte, se disponen de tal forma que la radiacin incida oblicuamente, recibiendo menos luz. Los cloroplastos se originan a partir de proplastidios, reaccin sta que es disparada por la luz, que provoca la diferenciacin del plastidio, apareciendo los pigmentos y la proliferacin de membranas, que origina los tilacoides y grana. As mismo, en el estroma del cloroplasto se encuentran pequeos pedazos circulares de ADN, dispuestos en doble hlice; parecidos al ADN de las mitocondrias y bacterias. El ADN del cloroplasto regula la sntesis del ARN ribosomal, del ARN de transferencia y de la Ribulosa 1,5 difosfato carboxilasa-oxigenasa (RUBISCO), enzima que cataliza la fijacin del CO2 en la fotosntesis. Sin embargo, la mayora de las protenas del cloroplasto, son sintetizadas en el citosol y transportadas al cloroplasto. Mitocondrias. Las clulas eucariticas poseen organelos complejos, denominados mitocondrias. Observadas con el microscopio ptico, se ven como pequeas esferas, bastones o filamentos, que varan en forma y tamao, comnmente miden de 0,5 a 1,0 de dimetro y de 1,0 a 4,0 de longitud. Son ms numerosas que los cloroplastos, pudindose encontrar hasta 1000 por clula, pero varias algas, incluyendo Chlorella tienen una sola por clula La mitocondria es el organelo responsable de la respiracin aerbica (que utiliza O2), un proceso en el cual un carbohidrato se oxida por completo en presencia de O2, convirtindose en CO2, H2 O y energa almacenada en forma de ATP. Las mitocondrias se dividen por fisin o biparticin, y todas se originan a partir de las mitocondrias contenidas en el zigoto; de tal forma que sus membranas no se derivan del sistema de endomembranas. Ellas contienen ADN circular y ribosomas pequeos (15 nm), en la matriz, de tal manera que son capaces de sintetizar algunas de sus propias protenas. Sin embargo, dependen tambin de protenas sintetizadas en el citoplasma que estn bajo el control nuclear.
Las mitocondrias tienen una doble membrana, la membrana externa es lisa, y acta como un colador, permitiendo el paso de muchas molculas pequeas; mientras que la membrana interna, muestra plegamientos denominados crestas, que aumentan la superficie interna. La membrana interna es selectivamente permeable, regulando el tipo de molculas que la atraviesan. El compartimiento interno encerrado por la membrana interna es la matriz, de naturaleza coloidal, que contiene las enzimas del ciclo de Krebs o del cido ctrico. En la membrana interna de las mitocondrias, se encuentran insertos los transportadores de electrones y la ATP sintetasa, realizndose en ella la fosforilacin oxidativa o sea la sntesis de ATP, acoplada al consumo de O2. Vacuolas. Son organelos caractersticos de las clulas vegetales, rodeados por una membrana denominada tonoplasto, que controla el transporte de solutos hacia adentro y hacia afuera de la vacuola; regulando el potencial hdrico de la clula a travs de la osmosis. La vacuola contiene iones inorgnicos, cidos orgnicos, azucares, enzimas, cristales de oxalato de calcio, y una variedad de metabolitos secundarios (alcaloides, taninos, ), que frecuentemente juegan un papel en la defensa de las plantas. Algunas vacuolas tienen altas concentraciones de pigmentos, hidrosolubles, que le dan la coloracin a muchas flores, hojas y a la raz de remolacha. Los colorantes vacuolares, de hojas y flores sirven para atraer los insectos que transportan el polen y, en parte funcionan como pigmentos protectores del exceso de radiacin. Las vacuolas pueden almacenar protenas, especialmente en legumbres y cereales, es importante sealar los granos de aleurona, en las clulas de la capa de aleurona de los cereales (trigo, cebada) o en los cotiledones de semillas de leguminosas (caraota, arveja, lenteja). Al germinar las semillas, las protenas son hidrolizadas y los aminocidos transferidos al embrin en crecimiento. Algunas vacuolas almacenan grasas como oleosomas o cuerpos grasos, p.ej. el endosperma del Ricinus communis (aceite de ricino). Las vacuolas son ricas en enzimas hidrolticas, como proteasas, ribonucleasas, y glicosidasas, que cuando se liberan en el citosol, participan en la degradacin celular durante la senescencia. Las vacuolas tienen un pH ms cido que el citosol, cualquier exceso de iones de hidrgeno en el citosol es bombeado hacia la vacuola, mantenindose la constancia del pH citoslico. En vista de la cantidad de sustancias que se acumulan en la vacuola, se ha pensado de ellas por mucho tiempo, que son como el botadero de productos de desechos celulares (sustancias ergsticas). Las vacuolas se originan a partir de pequeas vacuolas en clulas, jvenes, meristemticas del pice del tallo o de la raz, las que crecen con la clula, absorbiendo agua por osmosis y unindose unas con otras, hasta que se forman grandes vacuolas. Las pequeas vacuolas o provacuolas parecen formarse a partir del aparato de Golgi o del retculo endoplasmtico . El ncleo celular y sus componentes. El ncleo es el organelo celular ms conspicuo, tiene forma esfrica o globular, con un dimetro de 5 a 15 . Es el centro de control de la clula; sin embargo no es un organelo independiente, ya que debe obtener sus protenas del citoplasma. El ncleo contiene la mayor cantidad de ADN, al que se le da el nombre de genoma, est rodeado por una envoltura nuclear, compuesta de dos membranas, que se fusionan en algunos puntos formando poros nucleares, que permiten la comunicacin del interior del ncleo con el citoplasma celular. Pueden existir desde pocos a miles de poros en una envoltura nuclear. Algunas macromolculas del ncleo, incluyendo subunidades ribosomales, son capaces de atravesar los poros nucleares hacia el citosol y viceversa. El ncleo ejerce su control sobre las funciones celulares va ARNm (cido ribonucleico mensajero), determinando las enzimas que se fabrican en la clula y stas a su vez determinan las reacciones qumicas que se llevan a cabo, y por ende la estructura y funcin celular.
El ncleo es el sitio de almacenamiento y replicacin de los cromosomas, que estn compuestos de ADN y protenas acompaantes. El complejo ADN-protena (nucleoproteina), se denomina cromatina, que se observa dispersa durante la interfase. Aunque la cromatina pareciera estar desordenada, no es as; ya que est organizada en estructuras llamados cromosomas. La longitud de todo el ADN del genoma de una planta es millones de veces mayor que el dimetro del ncleo donde se encuentra, podemos establecer la analoga con una bola de hilo enrollada varios kilmetros de longitud, metida dentro de una pelota de golf. Cuando una clula se prepara para dividirse, el ADN y las protenas que forman cada cromosoma se enrollan ms estrechamente; los cromosomas se acortan, engruesan y se hacen visibles al microscopio. El ncleo contiene una solucin acuosa, repleta de enzimas, el nucleopasma, en el cual se encuentran suspendidos la cromatina o los cromosomas y los nuclolos. Como ya mencionamos, el ADN almacena informacin, en forma de genes, que son segmentos o secuencias de ADN que contienen toda la informacin gentica para originar un producto gnico determinado -ARN, protena-. El ncleo contiene uno o ms cuerpos esfricos (pueden ser hasta 4), los nuclolos, que pueden tener de 3 a 5 de dimetro. Los nuclolos son masas densas de fibras, de forma irregular, se tien de oscuro, que se encuentran suspendidos en el nucleoplasma. En ellos se pueden encontrar reas claras, llamadas vacuolas nucleolares, que son indicativos de un nuclolo muy activo. Las clulas meristemticas, generalmente tienen nuclolos ms grandes que las clulas maduras o latentes. En el nuclolo se fabrica el ARN ribosomal, que junto a las protenas sintetizadas en el citoplasma, forman los ribosomas. El ARN ribosomal es codificado por regiones especiales en los cromosomas denominadas regiones organizadoras del nuclolo. Los nucolos se observan bien durante la interfase de la mitosis, que es la fase de descanso de la divisin celular, pero cuando la clula comienza a dividirse, en la profase, desaparecen los nuclolos y la membrana nuclear, que se reabsorbe en el retculo endoplasmtico.
Вам также может понравиться
- Ficha Tarea 4-5 Biología 3°añoДокумент3 страницыFicha Tarea 4-5 Biología 3°añocarolinaОценок пока нет
- Biología Guía 1 Respuestas FFFДокумент6 страницBiología Guía 1 Respuestas FFFAlejandra AlejandraОценок пока нет
- Temario 1.1.5. Morfofuncion de Membrana Plasmatica, Citoplasma y Organelos ResueltoДокумент6 страницTemario 1.1.5. Morfofuncion de Membrana Plasmatica, Citoplasma y Organelos ResueltoAdrian Araya ChazarroОценок пока нет
- Unidad I Introduccion A La Fisiologia VegetalДокумент11 страницUnidad I Introduccion A La Fisiologia VegetalDeivis QuispeОценок пока нет
- Retículo Endoplasmático - Microtubulo - RibosomasДокумент4 страницыRetículo Endoplasmático - Microtubulo - RibosomasAriel CQОценок пока нет
- Biología 3año TP3Документ9 страницBiología 3año TP3Bravo NoeliaОценок пока нет
- Guía Biología Unidad Cero 2do MedioДокумент4 страницыGuía Biología Unidad Cero 2do MedioCris Tian Ignacio AstudilloОценок пока нет
- NOTAS DE LOS SEMINARIOS BIOLOGÍA 2 CBCДокумент146 страницNOTAS DE LOS SEMINARIOS BIOLOGÍA 2 CBCAxel MaturanoОценок пока нет
- Guia de Practica Histologia 2013Документ136 страницGuia de Practica Histologia 2013Gianella Peña MendozaОценок пока нет
- Material 2020 - 4to BiologiaДокумент16 страницMaterial 2020 - 4to BiologiaWilber Ardiles del ArroyoОценок пока нет
- CelulaДокумент20 страницCelulaEricka LizbethОценок пока нет
- Pau Criteris 2011-19 CastellanoДокумент70 страницPau Criteris 2011-19 CastellanoHugo Álvarez CanoОценок пока нет
- Guía de Ejercicios Tipo PSUДокумент2 страницыGuía de Ejercicios Tipo PSUTatianopolisОценок пока нет
- Aparato de Golgi y LisosomasДокумент16 страницAparato de Golgi y LisosomasOmar Dario Urrego CardenasОценок пока нет
- UntitledДокумент25 страницUntitledEfrain RochaОценок пока нет
- CelulaДокумент25 страницCelulaEdgar TintaОценок пока нет
- Producto Academico #02Документ11 страницProducto Academico #02UnPoCoDeToDo100% (1)
- BIOLOGÍAДокумент8 страницBIOLOGÍADiana CortesОценок пока нет
- CELULAДокумент10 страницCELULAMarioAlbertoAlejandreTorresОценок пока нет
- Organelos SubcelularesДокумент61 страницаOrganelos SubcelularesReinaldo Rodriguez Guerrero100% (4)
- CitoplasmaДокумент5 страницCitoplasmaUri GonzalezОценок пока нет
- Lactancia Materna - 4Документ67 страницLactancia Materna - 4Adrian Betanzos Rodriguez100% (2)
- Segundo Parcial Biologia General 2021 P1Документ175 страницSegundo Parcial Biologia General 2021 P1Monica RodriguezОценок пока нет
- BiologíaДокумент4 страницыBiologíaJuan MosqueraОценок пока нет
- Preguntas PAU NúcleoДокумент5 страницPreguntas PAU NúcleoPilar HidalgoОценок пока нет
- Diap Unidad 13 Apa Golgi-Liso-PeroxДокумент15 страницDiap Unidad 13 Apa Golgi-Liso-PeroxMaria MaciasОценок пока нет
- La CélulaДокумент10 страницLa Célulaheidi monestelОценок пока нет
- Aparato de GolgiДокумент3 страницыAparato de GolgiDiana Garcia PiñaОценок пока нет
- Trabajo Con Nota Primero Medio OrganelosДокумент3 страницыTrabajo Con Nota Primero Medio OrganelosRoxana Velásquez NúñezОценок пока нет
- Pato p1 ESДокумент18 страницPato p1 ESJuan PachecoОценок пока нет