Вам также может понравиться
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Determination of Suitable Thin Layer Drying Curve Model For Some Vegetables and FruitsДокумент10 страницDetermination of Suitable Thin Layer Drying Curve Model For Some Vegetables and FruitsLe Quoc TuanОценок пока нет
- Exploratory or Confirmatory Factor AnalysisДокумент17 страницExploratory or Confirmatory Factor AnalysisodaiissaОценок пока нет
- The Effect of Post Harvest Calcium Application On Tissue Calcium Concentration, Quality Attributes, Incidence of Flesh Browning and Cell Wall Physicochemical Aspects of Peach FruitsДокумент8 страницThe Effect of Post Harvest Calcium Application On Tissue Calcium Concentration, Quality Attributes, Incidence of Flesh Browning and Cell Wall Physicochemical Aspects of Peach FruitsLe Quoc TuanОценок пока нет
- Minitab GuideДокумент21 страницаMinitab GuideBlazieОценок пока нет
- 01 Food ScienceДокумент7 страниц01 Food ScienceLe Quoc TuanОценок пока нет
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (120)
- Exercises For Passive VoiceДокумент6 страницExercises For Passive VoicechondrokoukiОценок пока нет
- BMJNPH 2022 000477.fullДокумент18 страницBMJNPH 2022 000477.fullcatiatexОценок пока нет
- English Lum Xii Mars 2024 & KunciДокумент5 страницEnglish Lum Xii Mars 2024 & KunciZhye Anhae ShieyОценок пока нет
- Enzyme PPT 12 PDFДокумент42 страницыEnzyme PPT 12 PDFAnonymous wY9ogndU100% (1)
- Nursing Care Plan For Breast Cancer NCP PDFДокумент2 страницыNursing Care Plan For Breast Cancer NCP PDFMaina BarmanОценок пока нет
- Fruit and Vegetable Canning Business PlanДокумент67 страницFruit and Vegetable Canning Business Plankyaq001100% (1)
- University of Iowa Student, Faculty, and Staff Directory 1982-1983Документ261 страницаUniversity of Iowa Student, Faculty, and Staff Directory 1982-1983Richard GarrisonОценок пока нет
- Afk For LunchДокумент2 страницыAfk For LunchTristan TkNyarlathotep Jusola-SandersОценок пока нет
- Project of In-Plant Training On Avalon Cosmetics Pvt. LTDДокумент77 страницProject of In-Plant Training On Avalon Cosmetics Pvt. LTDRAHUL CHAUDHARYОценок пока нет
- Tashelhit Textbook 2011Документ268 страницTashelhit Textbook 2011Massaoud Bicanimen100% (2)
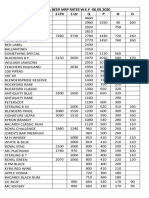
- Liquor & Beer MRP Rates Wef 17.12.2019 (1) - 1Документ2 страницыLiquor & Beer MRP Rates Wef 17.12.2019 (1) - 1Kondu Krishna Vamsi ChowdaryОценок пока нет
- All About ChocolateOДокумент23 страницыAll About ChocolateOMiae KimuОценок пока нет
- 12 The Malaysian Agricultural Industry in The New MillenniumДокумент20 страниц12 The Malaysian Agricultural Industry in The New MillenniumZainudin MohamedОценок пока нет
- Bahasa Inggris Kelas 2Документ3 страницыBahasa Inggris Kelas 2Fitriani Butterfly67% (3)
- Design Guide For HotelsДокумент33 страницыDesign Guide For HotelsSukhman Chawla100% (4)
- WSM West Salem MachineryДокумент7 страницWSM West Salem Machinerymostafa sherifОценок пока нет
- General Studies (Prelims) Paper - 1981Документ34 страницыGeneral Studies (Prelims) Paper - 1981suryakumar100% (1)
- New-Money FinalДокумент80 страницNew-Money FinalRashid LyonОценок пока нет
- Present Simple and Present ContinuousДокумент23 страницыPresent Simple and Present ContinuousLidia Gil González0% (1)
- TFJ1O Food Truck ProjectДокумент24 страницыTFJ1O Food Truck ProjectDivyasheni KrishnanОценок пока нет
- Pif 5 0Документ48 страницPif 5 0patricioacurioОценок пока нет
- RUSAYL TenantDirectory Eng 02-12-2022Документ41 страницаRUSAYL TenantDirectory Eng 02-12-2022Joseph AugustineОценок пока нет
- Finalized Training Report at Rich Life Dairies Limited 2Документ85 страницFinalized Training Report at Rich Life Dairies Limited 2asiri cloud100% (9)
- 12 Ways To Maximize Everyday in RamadanДокумент12 страниц12 Ways To Maximize Everyday in Ramadanthe_TRUTHОценок пока нет
- Food CombiningДокумент2 страницыFood Combiningmagda___Оценок пока нет
- Recipe For A Healthy Fruit Salad Activity CardДокумент4 страницыRecipe For A Healthy Fruit Salad Activity CardboobooОценок пока нет
- CVДокумент2 страницыCVJohn AttahОценок пока нет
- Detailed Lesson Plan in Tle Kitchen Utensils and EquipmentДокумент7 страницDetailed Lesson Plan in Tle Kitchen Utensils and EquipmentAprildhee Del Mundo Peña81% (88)
- Homework No. 2: GO Ian Patrick Q. April 9, 2020 BSPH 2AДокумент3 страницыHomework No. 2: GO Ian Patrick Q. April 9, 2020 BSPH 2AArman Renz PauloОценок пока нет
- Allrecipes - January 2015 USAДокумент122 страницыAllrecipes - January 2015 USALuana Burz100% (4)