Вам также может понравиться
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- Birth Plan + Hospital Bag Checklist: The Best EverДокумент32 страницыBirth Plan + Hospital Bag Checklist: The Best Everelly fadhliahОценок пока нет
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Mental Illness Attitude Scale (Modified OMICC QuestionnaireДокумент6 страницMental Illness Attitude Scale (Modified OMICC QuestionnaireSyed Faizan100% (7)
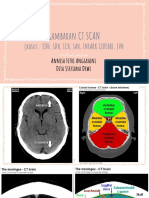
- Gambaran CT SCAN: (Kasus: Edh, SDH, Ich, Sah, Infark Cerebri, IvhДокумент44 страницыGambaran CT SCAN: (Kasus: Edh, SDH, Ich, Sah, Infark Cerebri, Ivhfahmi rosyadiОценок пока нет
- Empirical First Line AntibioticsДокумент1 страницаEmpirical First Line Antibioticsdiati zahrainiОценок пока нет
- Huntsville Businesses That Received More Than $150,000 From The Paycheck Protection ProgramДокумент14 страницHuntsville Businesses That Received More Than $150,000 From The Paycheck Protection ProgramJonathan GrassОценок пока нет
- Respiratory System Notes - KEYДокумент3 страницыRespiratory System Notes - KEYjoy100% (1)
- Journal Hydrodynamic Theory by Brnnstrm1986Документ5 страницJournal Hydrodynamic Theory by Brnnstrm1986Fifth TsuchikageОценок пока нет
- HIMSS EMRAM Stage 7 DMTДокумент20 страницHIMSS EMRAM Stage 7 DMTclrhoadesОценок пока нет
- Bioequivalence Comparison Between Two Different Formulations of Alverine AriqДокумент5 страницBioequivalence Comparison Between Two Different Formulations of Alverine AriqHendri WijayaОценок пока нет
- Grievously: News Item 9: EARTHQUAKE 2Документ2 страницыGrievously: News Item 9: EARTHQUAKE 2roxana_boraОценок пока нет
- Daftar Obat Slow Moving Dan Ed Rawat Inap Maret 2021Документ8 страницDaftar Obat Slow Moving Dan Ed Rawat Inap Maret 2021Vima LadipaОценок пока нет
- Project1 NewДокумент43 страницыProject1 NewSharad DholeОценок пока нет
- Legumes - AnOverview PDFДокумент6 страницLegumes - AnOverview PDFMayuri JagtapОценок пока нет
- Nursing Care Plan HydrocephalusДокумент7 страницNursing Care Plan HydrocephalusFarnii MarquezОценок пока нет
- Model Paper Set-B Medicinal Chemistry-I Bp402T: Section A MM 75 1. Attempt All Questions in Brief. 10 X 1 20Документ1 страницаModel Paper Set-B Medicinal Chemistry-I Bp402T: Section A MM 75 1. Attempt All Questions in Brief. 10 X 1 20SwarnA TamtaОценок пока нет
- SVCTДокумент70 страницSVCTSvct DelhiОценок пока нет
- Body Surface Area: Average Adult Dose: ExampleДокумент3 страницыBody Surface Area: Average Adult Dose: ExampleDefi OktafiaОценок пока нет
- Jamnagar Thesis 2004Документ12 страницJamnagar Thesis 2004Swagath NОценок пока нет
- 3.pleural Effusion Imaging - Overview, Radiography, Computed TomographyДокумент32 страницы3.pleural Effusion Imaging - Overview, Radiography, Computed TomographyAchmadRizaОценок пока нет
- What Type of Resistance Training Program Can Help Increase Muscle MassДокумент75 страницWhat Type of Resistance Training Program Can Help Increase Muscle Massyokimura dimaunahanОценок пока нет
- PTP1 - Reading Test 5Документ17 страницPTP1 - Reading Test 5Anh DucОценок пока нет
- CBT IdДокумент14 страницCBT IdNASC8Оценок пока нет
- Kerala PДокумент106 страницKerala PJeshiОценок пока нет
- Documentation: Siji M. SunnyДокумент5 страницDocumentation: Siji M. SunnySUNNY__SIJIОценок пока нет
- Modified Fluid Wax Impression For A Severely Resorbed Edentulous Mandibular RidgeДокумент4 страницыModified Fluid Wax Impression For A Severely Resorbed Edentulous Mandibular RidgeChepe LemusОценок пока нет
- Book Club Guide: INVINCIBLEДокумент1 страницаBook Club Guide: INVINCIBLEEpicReadsОценок пока нет
- Clinical Examination and Management of Impairments in VariousДокумент34 страницыClinical Examination and Management of Impairments in Variousmukesh pokharelОценок пока нет
- Science Blood TransfusionsДокумент10 страницScience Blood Transfusionsapi-237866019Оценок пока нет
- ADS ADEA ANZCA NZSSD - DKA - SGLT2i - Alert - Ver July 2022Документ3 страницыADS ADEA ANZCA NZSSD - DKA - SGLT2i - Alert - Ver July 2022tom.condon.02Оценок пока нет
- Kaloji Narayana Rao University Health Sciences College Allotment ListДокумент79 страницKaloji Narayana Rao University Health Sciences College Allotment ListVivek GuptaОценок пока нет