Вам также может понравиться
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- Tongue Thrust ExercisesДокумент18 страницTongue Thrust Exercisesfonomabel100% (1)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- CD Bernard LevinДокумент172 страницыCD Bernard LevinIndrani Das50% (2)
- Special Senses NotesДокумент17 страницSpecial Senses NotesizabelaОценок пока нет
- 14-Oral CavityДокумент36 страниц14-Oral CavitykavindukarunarathnaОценок пока нет
- The Function of TongueДокумент11 страницThe Function of TongueAndrea LawОценок пока нет
- Total Laryngectomy: Technique: Harvey M. Tucker, MDДокумент3 страницыTotal Laryngectomy: Technique: Harvey M. Tucker, MDKumaran Bagavathi RagavanОценок пока нет
- Tongue I LastДокумент30 страницTongue I LastEmuyeОценок пока нет
- Anatomy 10 YrsДокумент10 страницAnatomy 10 YrsAwantika RanjanОценок пока нет
- Thesis PDFДокумент43 страницыThesis PDFAbhishek PalОценок пока нет
- Lingual Pouch: DR - Srikanth GunturuДокумент18 страницLingual Pouch: DR - Srikanth GunturuSrikanth GunturuОценок пока нет
- 3 Case Report Geographic Tongue Haritsa BudimanДокумент10 страниц3 Case Report Geographic Tongue Haritsa Budimanharitsa budimanОценок пока нет
- Preview: Presbyphagia Markers in The Community-Dwelling Elderly PopulationДокумент13 страницPreview: Presbyphagia Markers in The Community-Dwelling Elderly PopulationvaleperoneОценок пока нет
- 1.4 Sense of Taste: Sense of Taste Detects The Presence of in Food & Drinks. Its Sensory Organ Is TheДокумент6 страниц1.4 Sense of Taste: Sense of Taste Detects The Presence of in Food & Drinks. Its Sensory Organ Is TheMissz WazeeОценок пока нет
- Digestion and Absorption: Digestive System of HumanДокумент4 страницыDigestion and Absorption: Digestive System of HumanrajaijahОценок пока нет
- Papillae of TongueДокумент28 страницPapillae of TongueDr. Janarthanan VОценок пока нет
- MCQ Final Oral Biology 2010Документ9 страницMCQ Final Oral Biology 2010Safura IjazОценок пока нет
- Articulatory Settings - Honikman (1964)Документ10 страницArticulatory Settings - Honikman (1964)Pronunciation ScienceОценок пока нет
- Oral Pathology - Tongue LesionДокумент26 страницOral Pathology - Tongue Lesionneji_murniОценок пока нет
- Armamentarium FOR Basic Oral Surgery: Deepthi P.R. Intern Dept. of Oral SurgeryДокумент140 страницArmamentarium FOR Basic Oral Surgery: Deepthi P.R. Intern Dept. of Oral SurgeryGabriela Cocieru MotelicaОценок пока нет
- Lingual Frenectomy Protocol Laser 2015Документ10 страницLingual Frenectomy Protocol Laser 2015jentotheskyОценок пока нет
- AnatomyДокумент91 страницаAnatomysarahОценок пока нет
- Nursing NotesДокумент6 страницNursing NotesAngel CauilanОценок пока нет
- Radya Agna Nugraha 1610211011Документ59 страницRadya Agna Nugraha 1610211011Radya AgnaОценок пока нет
- Developmental Disorders of Oral Cavity PDFДокумент121 страницаDevelopmental Disorders of Oral Cavity PDFMohammed Nabeel100% (1)
- Role of Tongue N Floor of Mouth in CD TRTДокумент18 страницRole of Tongue N Floor of Mouth in CD TRTashoorocksОценок пока нет
- Biopsy InterpretationДокумент474 страницыBiopsy InterpretationSandoz100% (1)
- лекция 8 Phlegmon of the floor of the mouth. Topographic anatomy of the floor of the mouth, sources of infection, possible ways of spreading the infection. Clinic, differential diagnostics. Surgery. PДокумент41 страницалекция 8 Phlegmon of the floor of the mouth. Topographic anatomy of the floor of the mouth, sources of infection, possible ways of spreading the infection. Clinic, differential diagnostics. Surgery. Ptalalelsherif92Оценок пока нет
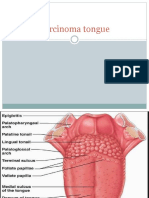
- Carcinoma TongueДокумент39 страницCarcinoma TongueVijay Krishna Murthy33% (3)
- Extraoral and Intraoral Exam: Dentalelle TutoringДокумент69 страницExtraoral and Intraoral Exam: Dentalelle TutoringEviОценок пока нет
- Compa Ana Digestive SystemДокумент12 страницCompa Ana Digestive SystemLemuel ValerioОценок пока нет