Вам также может понравиться
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (894)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (119)
- MethodsinMolecularBiology1200JunHirabayashieds. Lectins - MethodsandProtocols HumanaPress2014 PDFДокумент601 страницаMethodsinMolecularBiology1200JunHirabayashieds. Lectins - MethodsandProtocols HumanaPress2014 PDFSIVARAMANJAGANATHANОценок пока нет
- Genetic Re Combination and Its Molecular MechanismsДокумент20 страницGenetic Re Combination and Its Molecular MechanismsNabamita Baruah100% (2)
- 94Документ95 страниц94lthyaguОценок пока нет
- Conference AnnouncementДокумент1 страницаConference AnnouncementjchickОценок пока нет
- Microbiology SyllabusДокумент56 страницMicrobiology SyllabusYaswanth SinhaОценок пока нет
- Phylogenomics PDFДокумент227 страницPhylogenomics PDFalbertoadalidОценок пока нет
- Nucleic AcidsДокумент36 страницNucleic AcidsAman UllahОценок пока нет
- 6 DR PHAM Probio Feeds PDFДокумент27 страниц6 DR PHAM Probio Feeds PDFRinaldiОценок пока нет
- Klasifikasikan Berdasarkan Asalnya Stem CellДокумент2 страницыKlasifikasikan Berdasarkan Asalnya Stem CellfathinanisaОценок пока нет
- Exam blueprintДокумент2 страницыExam blueprintcoolchapОценок пока нет
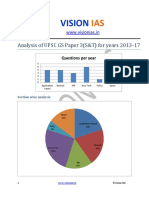
- Analysis of Upsc Gs Paper 3 Science and Tech Without ApproachДокумент2 страницыAnalysis of Upsc Gs Paper 3 Science and Tech Without ApproachJames KnotОценок пока нет
- Figure Mutant Paper 08222022Документ14 страницFigure Mutant Paper 08222022chau418Оценок пока нет
- Levine H-Challenges in The CGMP Manufacturing of Hescs-Lessons Learned From Monoclonal Antibodies-RprintДокумент24 страницыLevine H-Challenges in The CGMP Manufacturing of Hescs-Lessons Learned From Monoclonal Antibodies-RprintChilliG35Оценок пока нет
- Contrat - Technip - Sanofi - Algérie 2 PDFДокумент8 страницContrat - Technip - Sanofi - Algérie 2 PDFnadjib62Оценок пока нет
- Punnet QuizДокумент2 страницыPunnet Quizapi-351157612Оценок пока нет
- Notice: Reports and Guidance Documents Availability, Etc.: Q10 Pharmaceutical Quality SystemДокумент2 страницыNotice: Reports and Guidance Documents Availability, Etc.: Q10 Pharmaceutical Quality SystemJustia.comОценок пока нет
- Year II Biochemistry Department 1st SemesterTeaching Timetable NOV 2017 - MARCH 2018s-1Документ1 страницаYear II Biochemistry Department 1st SemesterTeaching Timetable NOV 2017 - MARCH 2018s-1Kevin ObureОценок пока нет
- RGNF SC 2016 17Документ251 страницаRGNF SC 2016 17Sukant MakhijaОценок пока нет
- USFDA Guidance For Industry - PSUR - What To ReportДокумент7 страницUSFDA Guidance For Industry - PSUR - What To ReportErshad Shafi AhmedОценок пока нет
- ID Regulasi Keamanan Hayati Produk Rekayasa PDFДокумент18 страницID Regulasi Keamanan Hayati Produk Rekayasa PDFPangihutan manaluОценок пока нет
- API SM Part 1Документ7 страницAPI SM Part 1Ronaldo JanglinОценок пока нет
- Endothelin-1 Gene Regulation: The FASEB JournalДокумент13 страницEndothelin-1 Gene Regulation: The FASEB JournalElena EllaОценок пока нет
- Programme - M.Sc. Biotechnology: L T P/S SW/F W Total Credit Units 3 0 0 0 3Документ4 страницыProgramme - M.Sc. Biotechnology: L T P/S SW/F W Total Credit Units 3 0 0 0 3thamizh555Оценок пока нет
- Biosimilares 2.0Документ8 страницBiosimilares 2.0Carlos Julio Nova LopezОценок пока нет
- Chapters 15Документ7 страницChapters 15bakelake101Оценок пока нет
- Bio in For MaticsДокумент17 страницBio in For MaticsSayan MandalОценок пока нет
- 08 Lac OPERONДокумент14 страниц08 Lac OPERONAlok SinghОценок пока нет
- The Golden Rice by Ambesh SrivastavaДокумент26 страницThe Golden Rice by Ambesh Srivastavascribd4986100% (1)
- Time and ConcentrationДокумент2 страницыTime and ConcentrationikrimakhaerunnisaОценок пока нет
- Genetic Code - Wikipedia DiscoveryДокумент2 страницыGenetic Code - Wikipedia Discoveryapi-243852896Оценок пока нет