Вам также может понравиться
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (119)
- Proposal Presentation Measurement of Amplitude of Accommodation in Different MethodsДокумент23 страницыProposal Presentation Measurement of Amplitude of Accommodation in Different MethodsTanzia HabibОценок пока нет
- Using Liquid Crystal Glasses To Treat Amblyopia in ChildrenДокумент3 страницыUsing Liquid Crystal Glasses To Treat Amblyopia in ChildrenWagner de AvizОценок пока нет
- Smartphone-based screening for diabetic retinopathy in IndiaДокумент10 страницSmartphone-based screening for diabetic retinopathy in IndiaVivi DeviyanaОценок пока нет
- Racgp Floaters and Flashes OpthalmologyДокумент3 страницыRacgp Floaters and Flashes OpthalmologyBeaulah HunidzariraОценок пока нет
- Delayed Visual Maturation in Infants: Andrew Tatham, Saurabh JainДокумент12 страницDelayed Visual Maturation in Infants: Andrew Tatham, Saurabh JainAffannul HakimОценок пока нет
- Senile Cataract Clinical Presentation and CausesДокумент4 страницыSenile Cataract Clinical Presentation and CausesAhmad FahroziОценок пока нет
- Igcse AccommodationДокумент3 страницыIgcse AccommodationChr1zFX4Оценок пока нет
- HPE Color Palette Asset Overview 050415Документ1 страницаHPE Color Palette Asset Overview 050415Henderson and SantosОценок пока нет
- OT - Grandjean - Ergonomics in Computerized Offices PDFДокумент238 страницOT - Grandjean - Ergonomics in Computerized Offices PDFAubrey Vale SagunОценок пока нет
- Anatomy of The EyeДокумент3 страницыAnatomy of The EyeNABEEL100% (1)
- Bionic EyeДокумент15 страницBionic EyeAravind RadhakrishnanОценок пока нет
- Fundus Examination PDFДокумент125 страницFundus Examination PDFAravind Kumar100% (5)
- Special SensesДокумент10 страницSpecial SensesEthel May AlabastroОценок пока нет
- Neuromuscular Physical TherapyДокумент13 страницNeuromuscular Physical Therapypearl042008100% (3)
- Amblyopia: Glomarie Hope A. Paquera Danica Ilah M. SincoДокумент39 страницAmblyopia: Glomarie Hope A. Paquera Danica Ilah M. SincoJan IrishОценок пока нет
- Elements of Design Tle 8Документ8 страницElements of Design Tle 8Arabelle MorilloОценок пока нет
- Spasm of AccommdationДокумент29 страницSpasm of Accommdationabdulaziz100% (1)
- Optometry AND Ophthalmology: Kerala Government Optometrists Association PSC TrainingДокумент137 страницOptometry AND Ophthalmology: Kerala Government Optometrists Association PSC TrainingHasan Ansari100% (1)
- PM Michaell Gazzaniga The Social BrainДокумент116 страницPM Michaell Gazzaniga The Social BrainVitor AraújoОценок пока нет
- Optics and RefractionДокумент135 страницOptics and RefractionAshraf Rapti100% (1)
- Colour: Art & Science - MollonДокумент25 страницColour: Art & Science - MollonTatiana MaiaОценок пока нет
- What Is Psychology 3rd Edition by Ellen E. Pastorino - Test BankДокумент50 страницWhat Is Psychology 3rd Edition by Ellen E. Pastorino - Test BankalishcathrinОценок пока нет
- The Other Amsler ChartsFINALДокумент20 страницThe Other Amsler ChartsFINALDanielle SangalangОценок пока нет
- Color Blindness, OR Color Vision DeficienciesДокумент11 страницColor Blindness, OR Color Vision DeficienciesSalih AliОценок пока нет
- Module f3 SC Chapter 1Документ26 страницModule f3 SC Chapter 1HackqRОценок пока нет
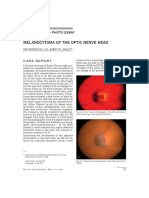
- Melanocytoma of The Optic Nerve Head: Clinical Case - Photo EssayДокумент5 страницMelanocytoma of The Optic Nerve Head: Clinical Case - Photo EssayKaran KumarswamyОценок пока нет
- Maryam PresentationДокумент9 страницMaryam PresentationUsman MuneerОценок пока нет
- Anatomy and Function of The EyeДокумент3 страницыAnatomy and Function of The EyeAljhon DelfinОценок пока нет
- Visual System ReportДокумент7 страницVisual System ReportAndre DueñasОценок пока нет
- Practical Refraction EnglishДокумент56 страницPractical Refraction Englisherin100% (2)