Вам также может понравиться
- Texto 9 Translocação de FotoassimiladosДокумент9 страницTexto 9 Translocação de FotoassimiladosBrunno SantosОценок пока нет
- TransolutoДокумент10 страницTransolutoRogerio Albino CanaeОценок пока нет
- Relatório Biologia - Transporte Nas Plantas PDFДокумент30 страницRelatório Biologia - Transporte Nas Plantas PDFO Sérgio ConceiçãoОценок пока нет
- Pemba UcmДокумент7 страницPemba UcmSardio Dos Santos SantosОценок пока нет
- Transl. Floema - Maria FernandaДокумент3 страницыTransl. Floema - Maria FernandaMaria FernandaОценок пока нет
- INTRODUÇÃO Transporte Do FloemaДокумент10 страницINTRODUÇÃO Transporte Do FloemaSérgio JerónimoОценок пока нет
- FloemaДокумент31 страницаFloemaLaís BarretoОценок пока нет
- Obtençao de Materia ResumoДокумент4 страницыObtençao de Materia Resumofrancisca soaresОценок пока нет
- Plasmodesmos Transporte Simplástico de Herbicidas Na PlantaДокумент10 страницPlasmodesmos Transporte Simplástico de Herbicidas Na Plantabaleiaj7Оценок пока нет
- BIOLOGIA RelatorioДокумент16 страницBIOLOGIA RelatorioAlexandra Ramos100% (2)
- A Obtenção de Matéria Pelos Seres HeterotróficosДокумент3 страницыA Obtenção de Matéria Pelos Seres HeterotróficosSara AlvesОценок пока нет
- 2020 1 ECE Transporte de FotoassimiladosДокумент16 страниц2020 1 ECE Transporte de FotoassimiladosManuela LopesОценок пока нет
- Biologia Fisiologia Vegetal 09 Floema TranslocacaoДокумент26 страницBiologia Fisiologia Vegetal 09 Floema TranslocacaoZedasCoves_Оценок пока нет
- Relatorio Transporte de H2o Na PlantaДокумент7 страницRelatorio Transporte de H2o Na PlantaMirian FreitasОценок пока нет
- Membrana e TransportesДокумент23 страницыMembrana e TransportesmatldeОценок пока нет
- SistematicaДокумент6 страницSistematicaeduardo_kadeshОценок пока нет
- Transporte FloemaДокумент50 страницTransporte FloemaHeitor O'Dwyer BisnettoОценок пока нет
- Biologia 10º - Distribuição de Matéria Nas PlantasДокумент6 страницBiologia 10º - Distribuição de Matéria Nas Plantasmilenerato2240Оценок пока нет
- Ae bg10 B PPT MembranaДокумент24 страницыAe bg10 B PPT MembranaSusana AlvesОценок пока нет
- Transporte Nas Plantas PDFДокумент6 страницTransporte Nas Plantas PDFGildaMarisaОценок пока нет
- BiológicaДокумент2 страницыBiológicaGoncalo FernandesОценок пока нет
- BIOLOGIAДокумент7 страницBIOLOGIATomásОценок пока нет
- Modulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Документ13 страницModulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Akiane FelixОценок пока нет
- Obtenção de Matéria (Resumo Online)Документ7 страницObtenção de Matéria (Resumo Online)Maria Abrantes TavaresОценок пока нет
- Transporte Nas PlantasДокумент14 страницTransporte Nas PlantasDulce SantosОценок пока нет
- Resumos Biologia 4ºtesteДокумент20 страницResumos Biologia 4ºtesteRafaela CoelhoОценок пока нет
- Transporte Nas PlantasДокумент19 страницTransporte Nas Plantas16736 Matilde Oliveira FernandesОценок пока нет
- Sebenta Bio I 3 UДокумент24 страницыSebenta Bio I 3 Upatriciaruth099Оценок пока нет
- Exercícos F. Vegetal de Todos Os TópicosДокумент36 страницExercícos F. Vegetal de Todos Os TópicosJefferson DavidОценок пока нет
- Distribuição de Matéria - RESUMOДокумент9 страницDistribuição de Matéria - RESUMOAnabela Rodrigues GonçalvesОценок пока нет
- Coradin, J. H. 2010 - (DISSERTAÇAO) PDFДокумент141 страницаCoradin, J. H. 2010 - (DISSERTAÇAO) PDFMarla SaldanhaОценок пока нет
- Biologia 10ºanoДокумент47 страницBiologia 10ºanocarinafish100% (2)
- A OBTENÇÃO DE MATÉRIA PELOS SERES HETEROTRÓFICOS - ResumosДокумент7 страницA OBTENÇÃO DE MATÉRIA PELOS SERES HETEROTRÓFICOS - ResumosSonjaОценок пока нет
- ObtençãodematériaresumoДокумент7 страницObtençãodematériaresumogentedanaveОценок пока нет
- O Transporte Nas PlantasДокумент63 страницыO Transporte Nas PlantasVascoCruzОценок пока нет
- Condução Das SeivasДокумент8 страницCondução Das SeivasEliseu Júlio MendesОценок пока нет
- O Transporte de Matéria Orgânica em Plantas - Artigo Científico PTДокумент5 страницO Transporte de Matéria Orgânica em Plantas - Artigo Científico PTTIAGO FERNANDES LOUÇÃO 13274Оценок пока нет
- Aula - Transporte de Água e Fotoassimildaos - COMPLETAДокумент34 страницыAula - Transporte de Água e Fotoassimildaos - COMPLETAotacide RosarioОценок пока нет
- Botânica GeralДокумент11 страницBotânica GeralAba lichakaОценок пока нет
- DistribuicaodamateriaДокумент15 страницDistribuicaodamateriaHugo MartinsОценок пока нет
- O Transporte Nos Animais Biologia 10ºanoДокумент17 страницO Transporte Nos Animais Biologia 10ºanoTito CunhaОценок пока нет
- 2 - Transporte Nas PlantasДокумент7 страниц2 - Transporte Nas PlantasnesОценок пока нет
- 3-Material Abs TranslocДокумент28 страниц3-Material Abs TranslocAndré Junior OgliariОценок пока нет
- Anexo XVI - Transporte No Xilema e FloemaДокумент7 страницAnexo XVI - Transporte No Xilema e FloemaJohn Little LightОценок пока нет
- Resumos BiologiaДокумент5 страницResumos BiologiaMara CostaОценок пока нет
- Introdução Ao LophotrochozoaДокумент14 страницIntrodução Ao LophotrochozoaDione Queiroz Amorim100% (1)
- 6 FloemaДокумент19 страниц6 FloemaFilipe BarreiroОценок пока нет
- Biofisica Do Sistema ExcretorДокумент8 страницBiofisica Do Sistema ExcretorYarinha GomezОценок пока нет
- Trocas Gasosas Nos Seres MulticelularesДокумент4 страницыTrocas Gasosas Nos Seres MulticelularesMariana DiasОценок пока нет
- Apontamentos Biologia Maioresde23Документ31 страницаApontamentos Biologia Maioresde23Pedro Póvoa OlyОценок пока нет
- Estudo DirigidoДокумент7 страницEstudo DirigidoMárcio Batista80% (5)
- Questões para Estudo e Reflexão I (Respostas)Документ24 страницыQuestões para Estudo e Reflexão I (Respostas)pri_pereirasОценок пока нет
- Modelo Roteiro de Estudos 13.01 3Документ5 страницModelo Roteiro de Estudos 13.01 3Mirna BatistaОценок пока нет
- BIOLOGIA - Distribuição Da Matéria 10ºДокумент5 страницBIOLOGIA - Distribuição Da Matéria 10ºRita CarreiraОценок пока нет
- Linhagens de Organismos Metazoários 1 Autor João Miguel de Matos NogueiraДокумент14 страницLinhagens de Organismos Metazoários 1 Autor João Miguel de Matos NogueiraJoão Victor FischerОценок пока нет
- Transporte Nas PlantasДокумент9 страницTransporte Nas PlantasAEESGPОценок пока нет
- Botânica estrutural: morfologia e anatomia de traqueófitasОт EverandBotânica estrutural: morfologia e anatomia de traqueófitasОценок пока нет
- Feijao KC Campos GoytacasesДокумент5 страницFeijao KC Campos GoytacasesRODRIGOSNPОценок пока нет
- A Cultura Do MilhoДокумент37 страницA Cultura Do MilhoBRUNINHOGUERREIRO100% (7)
- Feijao KC Campos GoytacasesДокумент5 страницFeijao KC Campos GoytacasesRODRIGOSNPОценок пока нет
- Algod Contr Biol Artrop Com Pre Dad OresДокумент19 страницAlgod Contr Biol Artrop Com Pre Dad OresRODRIGOSNPОценок пока нет
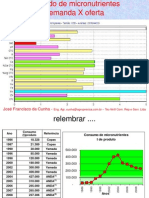
- 2-Franscisco Cunha Micronutrientes Nov-2010finalДокумент32 страницы2-Franscisco Cunha Micronutrientes Nov-2010finalRODRIGOSNPОценок пока нет
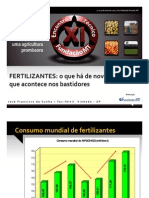
- Fertiliz Novidades - CunhaДокумент33 страницыFertiliz Novidades - CunhaRODRIGOSNPОценок пока нет
- Curso NeuroanatomiaДокумент49 страницCurso NeuroanatomiaJaqueline BoscoОценок пока нет
- tv5 em Ficha Avaliacao 6aДокумент3 страницыtv5 em Ficha Avaliacao 6aanitadiogoОценок пока нет
- Browning Do Tecido Adiposo Branco Lições de Modelos Experimentais - En.ptДокумент13 страницBrowning Do Tecido Adiposo Branco Lições de Modelos Experimentais - En.ptYara ZaninОценок пока нет
- Representantes Do Grupo Das Briófitas. As Samambaias São As Principais Representantes Do Grupo Das PteridófitasДокумент2 страницыRepresentantes Do Grupo Das Briófitas. As Samambaias São As Principais Representantes Do Grupo Das PteridófitasGuaracyara Ribas AugustoОценок пока нет
- Atlas de HematoДокумент34 страницыAtlas de HematoVitória MaiaОценок пока нет
- Apostila - Maratona Enem 2021 - Exercícios 3 MatériasДокумент114 страницApostila - Maratona Enem 2021 - Exercícios 3 MatériasAlinneCardosoОценок пока нет
- Ae bf10 Ban Exe 3Документ2 страницыAe bf10 Ban Exe 3Alexandra SoeiroОценок пока нет
- Aula 15 - Propagação de Plantas, Implantação e Manejo de ViveirosДокумент34 страницыAula 15 - Propagação de Plantas, Implantação e Manejo de Viveirosfernanda LimaОценок пока нет
- Cooper PeptideДокумент3 страницыCooper PeptideGustavo MüllerОценок пока нет
- Resenha - "Adaptation - Current Usages - Mary Jane West-Heberhard"Документ3 страницыResenha - "Adaptation - Current Usages - Mary Jane West-Heberhard"Eugenio FonsecaОценок пока нет
- Universidade Federal de Santa MariaДокумент52 страницыUniversidade Federal de Santa MariaGih DrenОценок пока нет
- Captura Híbrida para HPV de Alto E Baixo Risco: Leticia Oliveira Canario BelfortДокумент1 страницаCaptura Híbrida para HPV de Alto E Baixo Risco: Leticia Oliveira Canario BelfortLetícia BelfortОценок пока нет
- Protozoários, Bactérias e Vírus - 7 AnoДокумент7 страницProtozoários, Bactérias e Vírus - 7 AnoRayane CastroОценок пока нет
- Efeitos Biológicos Das Radiações IonizantesДокумент11 страницEfeitos Biológicos Das Radiações IonizantesAlberto Galdino SparrowОценок пока нет
- AULA 1 - Métodos de Estudo em PatologiaДокумент36 страницAULA 1 - Métodos de Estudo em PatologiaJoelia maiaОценок пока нет
- Introdução À Biologia ForenseДокумент50 страницIntrodução À Biologia ForenseAtaide VieiraОценок пока нет
- Prova 8° AnoДокумент3 страницыProva 8° AnoMaria ClaraОценок пока нет
- RPD Vol 12 #4 Dezembro 2017 Artigo Revisão Pág 159 167 PDFДокумент7 страницRPD Vol 12 #4 Dezembro 2017 Artigo Revisão Pág 159 167 PDFJulio cesarОценок пока нет
- Mecanismos de Herança - P2Документ5 страницMecanismos de Herança - P2Luana LouroОценок пока нет
- Ciclo de Vida Das Plantas Terrestres-1Документ8 страницCiclo de Vida Das Plantas Terrestres-1Agnaldo inoque diquissone Inoque diquissoneОценок пока нет
- Hematologia Basica Parte 3 - HematopoeseДокумент16 страницHematologia Basica Parte 3 - HematopoeseP2P3ESTATISTICA CIPTUR100% (1)
- Unid 1Документ23 страницыUnid 1renato bryanОценок пока нет
- 9969ef78 2021 REV Geral BOOKДокумент102 страницы9969ef78 2021 REV Geral BOOKRafael SchuenkОценок пока нет
- Lista de Exercicios OrganelasДокумент5 страницLista de Exercicios OrganelasCAMILA COELHO DE MELOОценок пока нет
- Ross Tecido EpitelialДокумент71 страницаRoss Tecido EpitelialMaria Eduarda SantosОценок пока нет
- Prova Célula Agro AДокумент2 страницыProva Célula Agro AAline Aparecida Bastos PortelaОценок пока нет
- 4 Questões Doenças Mieloproliferativas Crônicas-1Документ3 страницы4 Questões Doenças Mieloproliferativas Crônicas-1MatheusОценок пока нет
- Caderno de Exercicios VestibularesДокумент94 страницыCaderno de Exercicios VestibularesSteffany BarbosaОценок пока нет
- Soft Ticks of Cuba PDFДокумент208 страницSoft Ticks of Cuba PDFadsandorОценок пока нет
- Teoria Celular e Microscopia - ExercíciosДокумент5 страницTeoria Celular e Microscopia - ExercíciosMatheusОценок пока нет