Вам также может понравиться
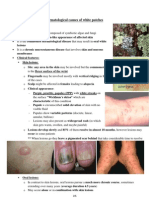
- 5 - Dermatological Causes of White PatchesДокумент6 страниц5 - Dermatological Causes of White PatchesRmz RabadiОценок пока нет
- Immuno - Lec 24Документ13 страницImmuno - Lec 24Rmz RabadiОценок пока нет
- Bonding and AdhesivesДокумент14 страницBonding and AdhesivesRmz RabadiОценок пока нет
- The Mid Exam Is Going To Be On The 4th of November at 11Документ11 страницThe Mid Exam Is Going To Be On The 4th of November at 11Rmz RabadiОценок пока нет
- Immuno - Lec 23Документ20 страницImmuno - Lec 23Rmz RabadiОценок пока нет
- The Mid Exam Is Going To Be On The 4th of November at 11Документ11 страницThe Mid Exam Is Going To Be On The 4th of November at 11Rmz RabadiОценок пока нет
- Immuno - Lec 22Документ11 страницImmuno - Lec 22Rmz RabadiОценок пока нет
- Immuno - Lec 21Документ13 страницImmuno - Lec 21Rmz RabadiОценок пока нет
- Immuno - Lec 19Документ13 страницImmuno - Lec 19Rmz RabadiОценок пока нет
- Immuno - Lec 20Документ13 страницImmuno - Lec 20Rmz RabadiОценок пока нет
- Immuno - Lec 17Документ18 страницImmuno - Lec 17Rmz RabadiОценок пока нет
- Immuno - Lec 18Документ21 страницаImmuno - Lec 18Rmz RabadiОценок пока нет
- Immuno - Lec 16Документ20 страницImmuno - Lec 16Rmz RabadiОценок пока нет
- Immuno - Lec 15Документ16 страницImmuno - Lec 15Rmz RabadiОценок пока нет
- Part One: Elastomers (Rubber Materials)Документ9 страницPart One: Elastomers (Rubber Materials)Rmz RabadiОценок пока нет
- RMZ Rabadi Dental AnatomyДокумент5 страницRMZ Rabadi Dental AnatomyRmz RabadiОценок пока нет
- GypsumДокумент12 страницGypsumRmz RabadiОценок пока нет
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- STSurg NumntДокумент11 страницSTSurg NumntrohiОценок пока нет
- Rotation Checklist - ClinicalДокумент3 страницыRotation Checklist - Clinicalapi-240994549100% (1)
- Finalpaper SchnakeДокумент6 страницFinalpaper Schnakeapi-315989347Оценок пока нет
- Case 29.07.09Документ51 страницаCase 29.07.09kanavgОценок пока нет
- BHU Daily Stock PerformaДокумент6 страницBHU Daily Stock PerformabilalОценок пока нет
- Final Brochure 2014Документ20 страницFinal Brochure 2014Annie NguyễnОценок пока нет
- Rubella: Congenital Clinically Significant Congenital MalformationsДокумент8 страницRubella: Congenital Clinically Significant Congenital MalformationsAlexandraОценок пока нет
- Philhealth Cf4 2017-2018Документ8 страницPhilhealth Cf4 2017-2018Julius Yves Dulfo BagacayОценок пока нет
- Trauma CounsellingДокумент2 страницыTrauma CounsellingSindujaa SelliahОценок пока нет
- Daftar Harga PT Bernofarm (RS Medirossa)Документ3 страницыDaftar Harga PT Bernofarm (RS Medirossa)Syaofi SofiОценок пока нет
- Momordica Cochinchinensis, Rosa Roxburghii, Wolfberry, and Sea Buckthorn-Highly Nutritional Fruits Supported by Tradition and ScienceДокумент8 страницMomordica Cochinchinensis, Rosa Roxburghii, Wolfberry, and Sea Buckthorn-Highly Nutritional Fruits Supported by Tradition and Sciencehaiquy174Оценок пока нет
- Iron Deficiency AnemiaДокумент25 страницIron Deficiency AnemiaDetria Rahma GastiОценок пока нет
- A 10-Point Plan For Avoiding Hyaluronic Acid Dermal Filler-Related Complications During Facial Aesthetic Procedures and Algorithms For ManagementДокумент9 страницA 10-Point Plan For Avoiding Hyaluronic Acid Dermal Filler-Related Complications During Facial Aesthetic Procedures and Algorithms For Managementleenatalia93100% (1)
- Postpartum Care: Emergency SignsДокумент13 страницPostpartum Care: Emergency SignsJohnnette BedoniaОценок пока нет
- Disaster Management Mental Health PerspectiveДокумент1 страницаDisaster Management Mental Health PerspectiveNatePrenticeОценок пока нет
- Effectiveness of A Desensitizing Agent Before In-Office Tooth Bleaching in Restored TeethДокумент7 страницEffectiveness of A Desensitizing Agent Before In-Office Tooth Bleaching in Restored TeethFer TorresОценок пока нет
- Ear DisordersДокумент10 страницEar DisordersPatzie MolinaОценок пока нет
- Mindful Eating and Dialectical Abstinence in DBT For Eating DisordersДокумент48 страницMindful Eating and Dialectical Abstinence in DBT For Eating DisordersArleta BluhmОценок пока нет
- Otitis MediaДокумент11 страницOtitis MediajessyОценок пока нет
- Chapter 5 - Counselling Skills: The GP As An Effective CounsellorДокумент19 страницChapter 5 - Counselling Skills: The GP As An Effective Counsellorprofarmah6150100% (2)
- Introduction To Burns: Bacteria VirusesДокумент6 страницIntroduction To Burns: Bacteria Virusesanon-50728Оценок пока нет
- Mind Mapping and Abdominal PainДокумент8 страницMind Mapping and Abdominal PaindtburrupОценок пока нет
- Pink Intimate Brochure PromoitaliaДокумент2 страницыPink Intimate Brochure PromoitaliaSkin365 Pty LtdОценок пока нет
- Ericksonian Hypnosis Techniques PDFДокумент2 страницыEricksonian Hypnosis Techniques PDFJakara25% (4)
- Cancer Pathophysiology FinalДокумент3 страницыCancer Pathophysiology FinalAngelique Ramos Pascua100% (1)
- PMLS Lesson 4Документ3 страницыPMLS Lesson 4Void MelromarcОценок пока нет
- Stugeron ForteДокумент4 страницыStugeron ForteJemsMei Comparativo MensuradoОценок пока нет
- Principles Involved in Bioassay by Different Methods A Minireview PDFДокумент18 страницPrinciples Involved in Bioassay by Different Methods A Minireview PDFPankaj KushwahОценок пока нет
- CKD CHCRTДокумент28 страницCKD CHCRTNurhidayati KeriyunОценок пока нет
- Preop ProofsДокумент14 страницPreop ProofsJanhvi AswaniОценок пока нет