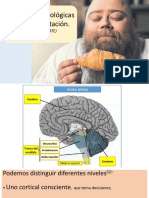
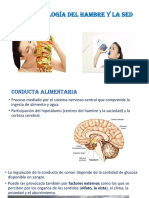
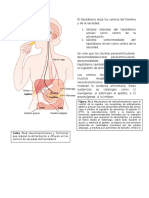
Вам также может понравиться
- Test TdahДокумент26 страницTest Tdahlissy_vija100% (10)
- Eje Cerebro IntestinoДокумент11 страницEje Cerebro IntestinoPatricia100% (2)
- Fisiologia Del HambreДокумент16 страницFisiologia Del HambrecaionevescaioОценок пока нет
- 3b Control Del Hambre y ApetitoДокумент36 страниц3b Control Del Hambre y ApetitoKarla MirellОценок пока нет
- Fisiología Del Hambre y SedДокумент15 страницFisiología Del Hambre y SedMiriam Mrn33% (3)
- Inteligencia Ejecutiva PDFДокумент13 страницInteligencia Ejecutiva PDFMarianneAS100% (1)
- Escala Australiana para Síndrome de AspergerДокумент3 страницыEscala Australiana para Síndrome de AspergerConstanza Vargas Barros100% (2)
- 2 Hambre, Apetito y SaciedadДокумент26 страниц2 Hambre, Apetito y SaciedadLuisB.CasasVargasОценок пока нет
- M2-T4 Regulación Fisiológica de La AlimentaciónДокумент10 страницM2-T4 Regulación Fisiológica de La AlimentaciónSil SerranoОценок пока нет
- Regulación de La IngestaДокумент3 страницыRegulación de La IngestaMaggie JacksonОценок пока нет
- Bases Biológicas Del Hambre y SedДокумент28 страницBases Biológicas Del Hambre y SedAbrahm Leon Chavez100% (1)
- Capacitación GPSДокумент40 страницCapacitación GPSYamil MarínОценок пока нет
- Hambre y SaciedadДокумент35 страницHambre y SaciedadMar OlaОценок пока нет
- Mecanismos Implicados en El Hambre y SaciedadДокумент4 страницыMecanismos Implicados en El Hambre y SaciedadDayraCampoverde50% (2)
- Cerdá - Avatares de Los AprendizajesДокумент281 страницаCerdá - Avatares de Los AprendizajesVanessa AybarОценок пока нет
- Zootecnia Alimentos y RacionamientoДокумент372 страницыZootecnia Alimentos y RacionamientoDon PepeОценок пока нет
- Fisiologia Del Hambre RevДокумент5 страницFisiologia Del Hambre RevcervantesОценок пока нет
- FISIOLOGIA DEL HAMBRE Y DEL APETITO. Documento #2Документ6 страницFISIOLOGIA DEL HAMBRE Y DEL APETITO. Documento #2Karla Elizabeth Fuentes LariosОценок пока нет
- 2 Hambre, Apetito y SaciedadДокумент23 страницы2 Hambre, Apetito y SaciedadCarlos Eduardo Layme MamaniОценок пока нет
- Metabolismo y Control Del CrecimientoДокумент30 страницMetabolismo y Control Del CrecimientoluciaslabochОценок пока нет
- HIPERFAGIAДокумент16 страницHIPERFAGIAKarol JaramilloОценок пока нет
- Fisiologia Hambre ApetitoДокумент60 страницFisiologia Hambre ApetitoArtur CrossОценок пока нет
- Resumen Mecanismos Neurobiologicos (Uno)Документ81 страницаResumen Mecanismos Neurobiologicos (Uno)fabiola alejandraОценок пока нет
- Neurofisiología Del Hambre y La SedДокумент15 страницNeurofisiología Del Hambre y La SedMitocondriaeriОценок пока нет
- Conducta Al Comer Estudia4Документ8 страницConducta Al Comer Estudia4marisol castilloОценок пока нет
- FORO-Respuestas de Preguntas Las OrientadorasДокумент2 страницыFORO-Respuestas de Preguntas Las OrientadorasMichael David Ramirez BayehОценок пока нет
- Procesos Biologicos Neuropsicologicos Unidad 5 (2021)Документ9 страницProcesos Biologicos Neuropsicologicos Unidad 5 (2021)Camila RadinoОценок пока нет
- ObesidadДокумент7 страницObesidadMaria NicoyaОценок пока нет
- Tema 2 PsicobioДокумент8 страницTema 2 PsicobioIrene Cepeda AlmagroОценок пока нет
- La Obesidad Es Una Enfermedad Crónica Que Abarca Muchos Factores y Que Se Caracteriza Por El Aumento Del Tejido Adiposo Que Ha Ido en Gran AumentoДокумент3 страницыLa Obesidad Es Una Enfermedad Crónica Que Abarca Muchos Factores y Que Se Caracteriza Por El Aumento Del Tejido Adiposo Que Ha Ido en Gran AumentoYessenia Rodriguez VivancoОценок пока нет
- El Apetito y La Saciedad - PPTX OficialДокумент23 страницыEl Apetito y La Saciedad - PPTX OficialSamuel Z Vega Miranda100% (2)
- Fc. - Tema 8Документ10 страницFc. - Tema 8Beatriz CastellanoОценок пока нет
- Hormonas Reguladoras Del HambreДокумент2 страницыHormonas Reguladoras Del Hambredianaale97Оценок пока нет
- DigestivoДокумент9 страницDigestivoAngélicaОценок пока нет
- Regulación de La IngestaДокумент8 страницRegulación de La IngestasireyerbcОценок пока нет
- Etapas de La Respiración y AnatomíaДокумент19 страницEtapas de La Respiración y AnatomíaAbigail MuzaОценок пока нет
- Regulación Central Del MetabolismoДокумент4 страницыRegulación Central Del Metabolismogrupo6 mktОценок пока нет
- Hormoonas Secretadas Por El IntestinoДокумент12 страницHormoonas Secretadas Por El IntestinoOlmos José EnriqueОценок пока нет
- Monografía Motivación y EmociónДокумент12 страницMonografía Motivación y EmociónGary KruzОценок пока нет
- FISIOLOGIADIGESTIVAДокумент9 страницFISIOLOGIADIGESTIVAToy CarbajalОценок пока нет
- Ensayo de 300 Palabras Sobre Las Funciones Principales de Los Núcleos Hipotalámicos Que Participan en La Ingesta AlimenticiaДокумент5 страницEnsayo de 300 Palabras Sobre Las Funciones Principales de Los Núcleos Hipotalámicos Que Participan en La Ingesta AlimenticiaBelinda CoelloОценок пока нет
- Afi2 U1 Ea DusmДокумент7 страницAfi2 U1 Ea DusmSardaneta SweetОценок пока нет
- HipotalamoДокумент2 страницыHipotalamodaniela escalonaОценок пока нет
- Neurociencias - Prueba 2Документ14 страницNeurociencias - Prueba 2Ignacia MarambioОценок пока нет
- Fisiologia Uabp7Документ5 страницFisiologia Uabp7Santiago SimonielloОценок пока нет
- Factores Del HambreДокумент13 страницFactores Del Hambrearelii martinezОценок пока нет
- Tema 3. Hambre y ApetitoДокумент10 страницTema 3. Hambre y ApetitoXabier TolarrajaОценок пока нет
- Notas Procesos Del Hambre y SaciedadДокумент2 страницыNotas Procesos Del Hambre y SaciedadJimena MJОценок пока нет
- Afi2 U1 Ea KatmДокумент6 страницAfi2 U1 Ea KatmKaty tmОценок пока нет
- Conducta Alimentaria Hambre y Sed.Документ5 страницConducta Alimentaria Hambre y Sed.LuisОценок пока нет
- PALIMENTARIA5Документ12 страницPALIMENTARIA5Lucía ArenasОценок пока нет
- Ansiedad y AlimentaciónДокумент15 страницAnsiedad y AlimentaciónMervin ChávezОценок пока нет
- Resumen, Tercer Corte (PSICOBIOLOGICAS)Документ9 страницResumen, Tercer Corte (PSICOBIOLOGICAS)lauraОценок пока нет
- 8 Octava Clase - Motivos Primarios y SecundariosДокумент23 страницы8 Octava Clase - Motivos Primarios y SecundariosAlexander Chura Limachi25% (4)
- Psicofisiologia 2Документ44 страницыPsicofisiologia 2Ruben Rodriguez GarzaОценок пока нет
- Afi2 U1 Ea MarzДокумент8 страницAfi2 U1 Ea Marzmarisol100% (1)
- Conducta MotivadaДокумент13 страницConducta MotivadaArlys Romero PlazaОценок пока нет
- Afi2 U1 A2 DoccДокумент6 страницAfi2 U1 A2 DoccDorisliliaОценок пока нет
- Cap 72 GuytonДокумент6 страницCap 72 GuytonRosario BarrientosОценок пока нет
- Fases de La DigestiónДокумент1 страницаFases de La DigestiónSilvana CevallosОценок пока нет
- Trabajo Final para Psicofisiología 1 (UASD)Документ39 страницTrabajo Final para Psicofisiología 1 (UASD)Enmanuel RodriguezОценок пока нет
- LateralДокумент6 страницLateralDaniel TibataОценок пока нет
- Fisiologia Del ApetitoДокумент3 страницыFisiologia Del Apetitojohanesquivelca0% (1)
- RESUMENДокумент2 страницыRESUMENPaola VélezОценок пока нет
- Linfangitis Ulcerativa en El CaballoДокумент5 страницLinfangitis Ulcerativa en El CaballoYamil MarínОценок пока нет
- Presentación de SaccharinaДокумент9 страницPresentación de SaccharinaYamil MarínОценок пока нет
- Tumores Melanóticos en CaballosДокумент3 страницыTumores Melanóticos en CaballosYamil MarínОценок пока нет
- Manejo No Quirúrgico de Los Sarcoides en El CaballoДокумент6 страницManejo No Quirúrgico de Los Sarcoides en El CaballoYamil MarínОценок пока нет
- Ficha RiegoДокумент2 страницыFicha RiegoChad MercerОценок пока нет
- Informe Nacional Sobre Sanidad e Incouidad - NicaraguaДокумент136 страницInforme Nacional Sobre Sanidad e Incouidad - NicaraguaYamil Marín100% (1)
- Biotecnologia de La ReproduccionДокумент280 страницBiotecnologia de La ReproduccionMartín Matías Pérez Zanón100% (4)
- La Ingeniería Genética y La Agricultura PDFДокумент26 страницLa Ingeniería Genética y La Agricultura PDFYamil MarínОценок пока нет
- Clase 2 CONTROL NEURONAL Y HORMONAL DE LA VOLEMIA Y LA PRESIÓN ARTERIAL PDFДокумент4 страницыClase 2 CONTROL NEURONAL Y HORMONAL DE LA VOLEMIA Y LA PRESIÓN ARTERIAL PDFYamil MarínОценок пока нет
- Gal 03 PDFДокумент6 страницGal 03 PDFYamil MarínОценок пока нет
- Algunas Plantas Tóxicas para El Ganado LecheroДокумент9 страницAlgunas Plantas Tóxicas para El Ganado LecheroYamil MarínОценок пока нет
- Minerales en Los Pastos de Centroam+®ricaДокумент7 страницMinerales en Los Pastos de Centroam+®ricaYamil MarínОценок пока нет
- Cria VacunaДокумент73 страницыCria VacunaYamil MarínОценок пока нет
- Malezas TóxicasДокумент85 страницMalezas TóxicasYamil MarínОценок пока нет
- Fractura de TibiaДокумент3 страницыFractura de TibiaYamil MarínОценок пока нет
- Principios de Producción de Vacunas VeterinariasДокумент17 страницPrincipios de Producción de Vacunas VeterinariasYamil MarínОценок пока нет
- Nutrientes en La DietaДокумент20 страницNutrientes en La DietaYamil MarínОценок пока нет
- Trastorno LímiteДокумент3 страницыTrastorno LímiteAriana Marycel Quispe QuispeОценок пока нет
- La Educación Artística Da Un Paso Al Frente-1Документ64 страницыLa Educación Artística Da Un Paso Al Frente-1Rosa Cubillo LópezОценок пока нет
- Alteraciones D El PensamientoДокумент152 страницыAlteraciones D El PensamientoVictor Gavidia CaceresОценок пока нет
- Act08 ANÁLISIS FUNCIONALДокумент6 страницAct08 ANÁLISIS FUNCIONALMiguel DuqueОценок пока нет
- E - BanduraДокумент23 страницыE - BanduraARIANNAОценок пока нет
- Monografia 1Документ8 страницMonografia 1Katherin Gisela Choque BelloОценок пока нет
- Neuropsicologia 4Документ4 страницыNeuropsicologia 4Rossemery PeraltaОценок пока нет
- Cerebrum 2015Документ13 страницCerebrum 2015Paula Gutierrez BurgoaОценок пока нет
- Empatía - ¿Por Qué Es Importante - Consulta de Psicología EpsibaДокумент23 страницыEmpatía - ¿Por Qué Es Importante - Consulta de Psicología EpsibaOlivia RojasОценок пока нет
- Lateralización Del LenguajeДокумент2 страницыLateralización Del LenguajeManchapanta Henrry JavierОценок пока нет
- Teoria Neurofisiologica Papers PDFДокумент5 страницTeoria Neurofisiologica Papers PDFJacqueline RamosОценок пока нет
- La Personalidad FДокумент4 страницыLa Personalidad FRoberto BelbereОценок пока нет
- Cuadro Comparativo EnfoquesДокумент7 страницCuadro Comparativo EnfoquesYazmin Tatiana RODRIGUEZ DAZAОценок пока нет
- Desarrollo Emocional de 0 A 6 MesesДокумент1 страницаDesarrollo Emocional de 0 A 6 MesesLizbeth Dominguez TОценок пока нет
- Cuestionario Bulbo RaquideoДокумент2 страницыCuestionario Bulbo RaquideoLinda Zapata Elguezabal100% (1)
- Tarea 4 Psicologia Del AprendizajeДокумент3 страницыTarea 4 Psicologia Del AprendizajeOcalys HicianoОценок пока нет
- PSICOPATOLOGIA Exploracion de Las Funciones MentalesДокумент20 страницPSICOPATOLOGIA Exploracion de Las Funciones MentalesDidier F. Barreto100% (1)
- Temario Por Cuatri Psico2Документ13 страницTemario Por Cuatri Psico2Lau BolsuaОценок пока нет
- Sólo La Vida Puede Conocer La VidaДокумент17 страницSólo La Vida Puede Conocer La VidaFrancisca Nevenka AlarcónОценок пока нет
- F20 PsicopatologíaДокумент2 страницыF20 PsicopatologíaRaúl EduardoОценок пока нет
- Taller Anatomia y Fisiologia Del CerebroДокумент10 страницTaller Anatomia y Fisiologia Del CerebroKAREN GARZON MORAОценок пока нет
- Teoría de Los Rasgos Equipo AzulДокумент17 страницTeoría de Los Rasgos Equipo AzulJeffryОценок пока нет
- TRASTORNO DE PERSONALIDAD PARANOIDE WordДокумент9 страницTRASTORNO DE PERSONALIDAD PARANOIDE WordJanny RojasОценок пока нет
- Corte 4 Psicologia PositivaДокумент4 страницыCorte 4 Psicologia PositivaNeil Alexander FigueredoОценок пока нет
- Tu Tipo de Personalidad A Traves Del Eneagrama PsicologicoДокумент4 страницыTu Tipo de Personalidad A Traves Del Eneagrama PsicologicoMarina MoranОценок пока нет
- Conceptos DHPДокумент3 страницыConceptos DHPShanti LiОценок пока нет