Вам также может понравиться
- Measuring Health and Disease (Session 7 and 8) 2Документ37 страницMeasuring Health and Disease (Session 7 and 8) 2teklay100% (1)
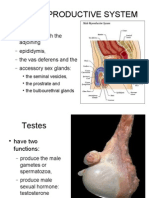
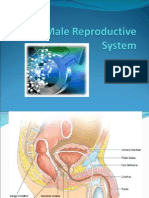
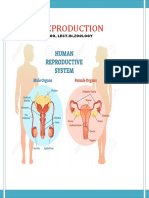
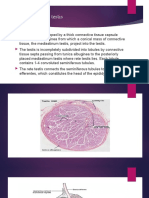
- The Male Genital SystemДокумент5 страницThe Male Genital SystemDr-AymanBahgatОценок пока нет
- The Male Genital SystemДокумент20 страницThe Male Genital SystemLittleChenaОценок пока нет
- MRS LabДокумент6 страницMRS Lab2012000600Оценок пока нет
- Male Reproductive Physiology 2021Документ188 страницMale Reproductive Physiology 2021Elijah Kihiu18Оценок пока нет
- Hist MaleДокумент7 страницHist MalezaidОценок пока нет
- Department of Morphology: Penza State University Medical InstituteДокумент18 страницDepartment of Morphology: Penza State University Medical InstituteunnatiОценок пока нет
- Reproductive System NotesДокумент12 страницReproductive System NotesTanaya PujareОценок пока нет
- Scrotum and OthersДокумент10 страницScrotum and OthersAmbg GhalyОценок пока нет
- Male Reproductive SystemДокумент16 страницMale Reproductive Systemqty9jgkpnzОценок пока нет
- Anatomy of The Prostate GlandДокумент18 страницAnatomy of The Prostate GlandRao Ghayoor DxОценок пока нет
- Anatomy and Physiology of Male Reproductive SystemДокумент52 страницыAnatomy and Physiology of Male Reproductive SystemAjay DОценок пока нет
- Reproductive SystemДокумент7 страницReproductive SystemElaine Eloisa CapilloОценок пока нет
- The Reproductive Systems AnatomyДокумент3 страницыThe Reproductive Systems AnatomyLilian JerotichОценок пока нет
- Malereproductive (Author T.globa)Документ40 страницMalereproductive (Author T.globa)Cristina caimac100% (1)
- Reproductive SystemДокумент42 страницыReproductive SystemAileen Grace Delima100% (1)
- Male R.S. Dr Hekmat هاند اوتДокумент15 страницMale R.S. Dr Hekmat هاند اوتJo adamОценок пока нет
- Lab Ex1-2 and Chick OrganogenesisiДокумент20 страницLab Ex1-2 and Chick OrganogenesisiLardel CarayОценок пока нет
- Med Lec Male Reproductive SystemДокумент67 страницMed Lec Male Reproductive Systemmiguel cuevasОценок пока нет
- Male and Female Genital Systems 2Документ30 страницMale and Female Genital Systems 2negmm2226Оценок пока нет
- Reproductive Systems A. B. C. DДокумент6 страницReproductive Systems A. B. C. DJamie BagundolОценок пока нет
- Human Reproduction and Reproductive HealthДокумент16 страницHuman Reproduction and Reproductive HealthHemanth GowdaОценок пока нет
- Reproductive System 1Документ9 страницReproductive System 1Marvel NnaemekaОценок пока нет
- Importance of Human Reproductive SystemДокумент3 страницыImportance of Human Reproductive Systemshn mzkОценок пока нет
- Chapter 26 M&Freprod 2304Документ55 страницChapter 26 M&Freprod 2304sairahhannahОценок пока нет
- 11 MaleДокумент14 страниц11 MaleJasmine KaurОценок пока нет
- Male Histology Trans-Batch 2023Документ11 страницMale Histology Trans-Batch 2023Hanako Sasaki AranillaОценок пока нет
- Lecture Notes Module - 3 - Male Reproductive Anatomy and PhysiologyДокумент8 страницLecture Notes Module - 3 - Male Reproductive Anatomy and PhysiologyMariaImranОценок пока нет
- Nurs 5343 Emergency Obstetrics CareДокумент92 страницыNurs 5343 Emergency Obstetrics CareRachel BiokoОценок пока нет
- Anatomy of The Male Reproductive SystemДокумент17 страницAnatomy of The Male Reproductive SystemFarhan haiderОценок пока нет
- GilsiuДокумент17 страницGilsiuexpostofaria78Оценок пока нет
- Male ReproductivehistoДокумент9 страницMale ReproductivehistoaikaОценок пока нет
- Maternal and ChildДокумент10 страницMaternal and Childryanescueta090702Оценок пока нет
- Male Reproductive SystemДокумент49 страницMale Reproductive SystemRinkish Dalliah50% (2)
- The Genital Organs DifferentiationДокумент31 страницаThe Genital Organs DifferentiationIdris LubisОценок пока нет
- Human ReproductionДокумент15 страницHuman ReproductionAnkit PruthwirajОценок пока нет
- Male Reproductive SystemДокумент6 страницMale Reproductive SystemCurex QAОценок пока нет
- Chapter 21 The Male Reproductive SystemДокумент6 страницChapter 21 The Male Reproductive SystemEllä Pabustan100% (1)
- Lab. NO. 4: System: ReproductiveДокумент10 страницLab. NO. 4: System: ReproductiveKarrar BassimОценок пока нет
- Structure of The TestisДокумент24 страницыStructure of The TestisBassam MohamedОценок пока нет
- Chapter: 3. Human Reproduction: The Male Reproductive SystemДокумент12 страницChapter: 3. Human Reproduction: The Male Reproductive Systemlogi nageshОценок пока нет
- 2776NEETZoologyReproductiveSystemTheory - Unlocked Opt PDFДокумент21 страница2776NEETZoologyReproductiveSystemTheory - Unlocked Opt PDFMd Rizwan AhmadОценок пока нет
- The Reproductive SystemДокумент68 страницThe Reproductive SystemSamuelОценок пока нет
- Male Reproduction System - Histology LectureДокумент28 страницMale Reproduction System - Histology LectureAstri Novia RizqiОценок пока нет
- Male Reproductive System: Nur AnisahДокумент33 страницыMale Reproductive System: Nur Anisahdhiet54Оценок пока нет
- Chapter 3 Human ReproductionДокумент13 страницChapter 3 Human ReproductionvidushivbpsОценок пока нет
- LECTURE 6 Male Reproductive System - Histology HB II 2023Документ76 страницLECTURE 6 Male Reproductive System - Histology HB II 2023Emmanuel AssopiahОценок пока нет
- Anatomy and Physiology of Male Reproductive SystemДокумент8 страницAnatomy and Physiology of Male Reproductive SystemAdor AbuanОценок пока нет
- Male ReproductiveДокумент59 страницMale ReproductiveChandan ShahОценок пока нет
- Histology of TestisДокумент2 страницыHistology of TestisyolandadwiooОценок пока нет
- Human ReproductionДокумент26 страницHuman Reproductionbarunmahto001Оценок пока нет
- Physics Class 10 ICSEДокумент5 страницPhysics Class 10 ICSEmohammedumar7864521Оценок пока нет
- Week 1 - Study QuestionsДокумент13 страницWeek 1 - Study Questionsjess waldenОценок пока нет
- CYTOLOGYДокумент16 страницCYTOLOGYRheimon Jay Abuan BalcitaОценок пока нет
- HAPP Supplementary Notes - Reproductive SystemДокумент15 страницHAPP Supplementary Notes - Reproductive SystemZuhri PorzaОценок пока нет
- Histology of The Male Reproductive SystemДокумент18 страницHistology of The Male Reproductive SystemMuhammad JameelОценок пока нет
- Xii Zool Ch3 Human Reproduction HssliveДокумент8 страницXii Zool Ch3 Human Reproduction HsslivePrituОценок пока нет
- Vpy 606 SeminarДокумент67 страницVpy 606 SeminarMohd Zakiuddin SiddiquiОценок пока нет
- 2080 Reproductive System of Frog Grade XIДокумент1 страница2080 Reproductive System of Frog Grade XIkunalkumaryadav086Оценок пока нет
- CHAPTER 4 Environmental EpidemiologyДокумент23 страницыCHAPTER 4 Environmental EpidemiologyteklayОценок пока нет
- Epidemiology For MPH StudentsДокумент91 страницаEpidemiology For MPH StudentsteklayОценок пока нет
- CHAPTER 6 Epidemiological SurveillanceДокумент50 страницCHAPTER 6 Epidemiological Surveillanceteklay100% (1)
- CHAPTER 2 Infectious EpidemiologyДокумент89 страницCHAPTER 2 Infectious EpidemiologyteklayОценок пока нет
- CHAPTER 3 Non-Communicable Disease EpidemiologyДокумент24 страницыCHAPTER 3 Non-Communicable Disease EpidemiologyteklayОценок пока нет
- Emerging and Re Emerging DiseasesДокумент49 страницEmerging and Re Emerging DiseasesteklayОценок пока нет
- 1 Problem Solving & Dicision MakingДокумент55 страниц1 Problem Solving & Dicision MakingteklayОценок пока нет
- 2 Management of Specialized Projects, ProgramesДокумент67 страниц2 Management of Specialized Projects, ProgramesteklayОценок пока нет
- Global Cooperation in Health: Manay KifleДокумент45 страницGlobal Cooperation in Health: Manay KifleteklayОценок пока нет
- Evidence-Based Public HealthДокумент21 страницаEvidence-Based Public HealthteklayОценок пока нет
- ch-8 Conflict Management in The Health Sector (2016 - 03 - 30 15 - 23 - 50 UTC)Документ61 страницаch-8 Conflict Management in The Health Sector (2016 - 03 - 30 15 - 23 - 50 UTC)teklayОценок пока нет
- Chapter-4 Health Professionals Motivation: Zemichael WGДокумент37 страницChapter-4 Health Professionals Motivation: Zemichael WGteklayОценок пока нет
- Ch-4 Leadership For MPHДокумент69 страницCh-4 Leadership For MPHteklayОценок пока нет
- CH-1 Introduction To HSM For MPHДокумент96 страницCH-1 Introduction To HSM For MPHteklayОценок пока нет
- L - 3 Outbreak InvestigationДокумент57 страницL - 3 Outbreak InvestigationteklayОценок пока нет
- Epidemics: Fessahaye Alemseged (MD, MPHE)Документ34 страницыEpidemics: Fessahaye Alemseged (MD, MPHE)teklay100% (1)
- L - 1measure of AssociationsДокумент48 страницL - 1measure of AssociationsteklayОценок пока нет
- Functions of Public HealthДокумент30 страницFunctions of Public Healthteklay100% (1)
- Course Syllabus of IT PG PDFДокумент39 страницCourse Syllabus of IT PG PDFteklayОценок пока нет
- Statistical Methods in Epi 2019Документ30 страницStatistical Methods in Epi 2019teklayОценок пока нет
- Validity of Epi Studies 2019Документ26 страницValidity of Epi Studies 2019teklayОценок пока нет
- Economic Evaluation (MPH)Документ52 страницыEconomic Evaluation (MPH)teklayОценок пока нет
- Screening: Berhe Beyene (Email:)Документ71 страницаScreening: Berhe Beyene (Email:)teklayОценок пока нет
- Aksum University-Department of Nursing For 3 Yr BSC Nursing StudentsДокумент21 страницаAksum University-Department of Nursing For 3 Yr BSC Nursing StudentsteklayОценок пока нет
- UTS ModuleДокумент62 страницыUTS ModuleMariane Joy TecsonОценок пока нет
- How Do Organisms ReproduceДокумент25 страницHow Do Organisms ReproduceTalent Public SchoolОценок пока нет
- Biology Study Material - Xii - 22-23 - 1Документ216 страницBiology Study Material - Xii - 22-23 - 1kalinayak.prasadОценок пока нет
- ReproДокумент80 страницReprocream o100% (1)
- The Human Reproductive System FinalДокумент16 страницThe Human Reproductive System FinalIlac Tristan BernardoОценок пока нет
- Development Reproductive SystemsДокумент59 страницDevelopment Reproductive SystemsAli AbdElnaby SalimОценок пока нет
- Diagestive, Urinary and Reproductive SystemДокумент39 страницDiagestive, Urinary and Reproductive SystemCarolina Calle HerreraОценок пока нет
- Las Science 5 Melc 1 q2 Week1Документ10 страницLas Science 5 Melc 1 q2 Week1Marjun BartoloОценок пока нет
- Class12th HUMAN-REPRODUCTION PDFДокумент29 страницClass12th HUMAN-REPRODUCTION PDFShubhang CharantimathОценок пока нет
- Gender Role SocializationДокумент67 страницGender Role SocializationJean Lyca Mea GerarcasОценок пока нет
- Lesson Plan Science 5Документ9 страницLesson Plan Science 5Marj Reña LunaОценок пока нет
- Applied Veterinary Andrology and Artificial InseminationДокумент146 страницApplied Veterinary Andrology and Artificial InseminationDeep Patel50% (2)
- 2.the Nursing Role in Reproductive and Sexual HealthДокумент70 страниц2.the Nursing Role in Reproductive and Sexual HealthAngel Gabriel FornillosОценок пока нет
- S10LT IIIb 34.1Документ4 страницыS10LT IIIb 34.1Marvi ValdezОценок пока нет
- 1 - Male Reproductive SystemДокумент10 страниц1 - Male Reproductive SystemIbsa AbdulwabОценок пока нет
- Reproductive SystemДокумент219 страницReproductive Systemtzushka_vipОценок пока нет
- Unit # 14 Bio Class 10Документ29 страницUnit # 14 Bio Class 10Muhammad Owais FayazОценок пока нет
- Sci10 Quarter 3 Module 1 NO ANSWER KEYДокумент30 страницSci10 Quarter 3 Module 1 NO ANSWER KEYNiña Miles TabañagОценок пока нет
- Probni Test 1. Godina - Ina KlipaДокумент4 страницыProbni Test 1. Godina - Ina KlipaMickoОценок пока нет
- Class 12 Biology Pock BookДокумент53 страницыClass 12 Biology Pock Bookbs.3136 VVS.Оценок пока нет
- Class 11 Concise Biology Notes For KVPY, NSEBДокумент121 страницаClass 11 Concise Biology Notes For KVPY, NSEBNischal Vir MОценок пока нет
- Sexual Reproduction Part 1Документ20 страницSexual Reproduction Part 1Jurez N. EvansОценок пока нет
- Assignment-The Male Reproductive SystemДокумент25 страницAssignment-The Male Reproductive Systemgopi_marco100% (1)
- Science V 2nd Quarter Lesson PlanДокумент154 страницыScience V 2nd Quarter Lesson PlanRHODA ESCOLANO100% (1)
- Human Reproductive SystemДокумент10 страницHuman Reproductive Systembryan100% (1)
- Disease Definisi Etiologi Manfes/findingДокумент3 страницыDisease Definisi Etiologi Manfes/findingRiani KusumadewiОценок пока нет
- EED 5 Unit 5Документ19 страницEED 5 Unit 5Lara Mariz FragataОценок пока нет
- Reviewer For RLE 101Документ9 страницReviewer For RLE 101ZLKASMdlasmОценок пока нет
- Human Sexuality Unit: Reproductive SystemsДокумент38 страницHuman Sexuality Unit: Reproductive SystemsMary Grace CacalОценок пока нет
- Assessing Children - Infancy Through Adolescence 1Документ118 страницAssessing Children - Infancy Through Adolescence 1kûrñï såñskrùthîОценок пока нет