Вам также может понравиться
- Clase03 Auscultacion Cardiaca I-Ii PDFДокумент25 страницClase03 Auscultacion Cardiaca I-Ii PDFDimitry Antonov Tartarov PupanovОценок пока нет
- Clase 2 Ciclo CardiacoДокумент14 страницClase 2 Ciclo CardiacoDimitry Antonov Tartarov PupanovОценок пока нет
- Arco Reflejo Placa MioneuralДокумент26 страницArco Reflejo Placa MioneuralDimitry Antonov Tartarov PupanovОценок пока нет
- Clase02 Semiologia CardioДокумент97 страницClase02 Semiologia CardioGoku Naruto VeguetaОценок пока нет
- Práctica N°9-Dr. Raúl Diaz - Matriz de Monitoreo, Supervisión y EvaluaciónДокумент9 страницPráctica N°9-Dr. Raúl Diaz - Matriz de Monitoreo, Supervisión y EvaluaciónDimitry Antonov Tartarov Pupanov100% (2)
- Practica Semana 4 - Casos Clínicos PDFДокумент27 страницPractica Semana 4 - Casos Clínicos PDFDimitry Antonov Tartarov PupanovОценок пока нет
- Sistema Endocrino Historia Clinica 2019Документ49 страницSistema Endocrino Historia Clinica 2019Dimitry Antonov Tartarov PupanovОценок пока нет
- Exposicion Liquidos CorporalesДокумент31 страницаExposicion Liquidos CorporalesDimitry Antonov Tartarov PupanovОценок пока нет
- HC 16 - Julián ArceДокумент13 страницHC 16 - Julián ArceDimitry Antonov Tartarov PupanovОценок пока нет
- Radiologia SNCДокумент52 страницыRadiologia SNCDimitry Antonov Tartarov PupanovОценок пока нет
- Fergus Fall Behaviour ScaleДокумент4 страницыFergus Fall Behaviour ScaleDimitry Antonov Tartarov Pupanov0% (1)
- Marcadores TumoralesДокумент30 страницMarcadores TumoralesDimitry Antonov Tartarov PupanovОценок пока нет
- Sapito SaponДокумент6 страницSapito SaponDimitry Antonov Tartarov PupanovОценок пока нет
- Test de Clasificación de Cartas deДокумент12 страницTest de Clasificación de Cartas deDimitry Antonov Tartarov PupanovОценок пока нет
- Tejido HematicoДокумент42 страницыTejido HematicoManuel CruzОценок пока нет
- El Esputo OsoДокумент5 страницEl Esputo OsoJalmer David DamnОценок пока нет
- ALERTA 10-12 DronedaronaДокумент1 страницаALERTA 10-12 DronedaronaMelinda FrederickОценок пока нет
- Taller #4Документ3 страницыTaller #4KRISS CONTRERASОценок пока нет
- Sistema DigestivoДокумент5 страницSistema DigestivoDeeyyn AicragОценок пока нет
- EmbriologiaДокумент5 страницEmbriologiaUrielОценок пока нет
- GPC Comp Linfoma PDFДокумент342 страницыGPC Comp Linfoma PDFluisaminelaОценок пока нет
- El Ajo Un Ingrediente SaludableДокумент3 страницыEl Ajo Un Ingrediente SaludableKirk Ldk67% (3)
- La Nutrición Uagrm-FinorДокумент29 страницLa Nutrición Uagrm-FinorMatthew BrooksОценок пока нет
- Cuadro Comparativo Teoria de La PersonalidadДокумент2 страницыCuadro Comparativo Teoria de La PersonalidadGuillermina Soria100% (1)
- Síntesis Del ColágenoДокумент4 страницыSíntesis Del ColágenoMaria Jose GonzalezОценок пока нет
- Apgar FamiliarДокумент4 страницыApgar FamiliarRonaldo_cseОценок пока нет
- Prospecto Urzac I0058-4Документ1 страницаProspecto Urzac I0058-4Harold83Оценок пока нет
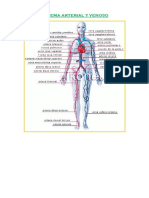
- Sistema Arterial y VenosoДокумент12 страницSistema Arterial y Venosoesthefanie ñontolОценок пока нет
- COLERAДокумент23 страницыCOLERAPedro PeraltaОценок пока нет
- Proceso de Atención en Enfermería Pae EjemploДокумент41 страницаProceso de Atención en Enfermería Pae EjemploJhoanaCarolinaRomeroRodriguezОценок пока нет
- 2 Fisiología Del Crecimiento FetalДокумент9 страниц2 Fisiología Del Crecimiento FetalSarahí SosaОценок пока нет
- EnanismoДокумент19 страницEnanismoShiri Angel ConradОценок пока нет
- Cuestionario de Micobacterias TuberculosisДокумент12 страницCuestionario de Micobacterias TuberculosisOrbely Garcia100% (2)
- Contaminacion de Los AlimentosДокумент15 страницContaminacion de Los Alimentosdeisi perales100% (2)
- Libro de Hemoterapia y Banco de Sangre 2014Документ28 страницLibro de Hemoterapia y Banco de Sangre 2014Guisela Mendocilla RamirezОценок пока нет
- AD2 MCD Manual (Spanish) 3-7-11Документ17 страницAD2 MCD Manual (Spanish) 3-7-11Jerome HickmanОценок пока нет
- Derrame PleuralДокумент26 страницDerrame PleuralJuan IgnacioОценок пока нет
- Clasificacion AOДокумент98 страницClasificacion AOEduardo Alexis Escoto LopezОценок пока нет
- GlucosidasaДокумент126 страницGlucosidasaMaritsa PerHerОценок пока нет
- Dependencia A Las Drogas, ¿UNA ENFERMEDAD CRONICA?Документ4 страницыDependencia A Las Drogas, ¿UNA ENFERMEDAD CRONICA?ariana_calderonlОценок пока нет
- Sindrome de TouretteДокумент25 страницSindrome de TouretteKaren PulidoОценок пока нет
- Fungi Cid AsДокумент8 страницFungi Cid AsYvonne RoLauОценок пока нет
- LeucemiasДокумент49 страницLeucemiasXavier AlexanderОценок пока нет
- Anemia Ferropenica 2Документ16 страницAnemia Ferropenica 2Bella Stefanny Morales GuevaraОценок пока нет