Вам также может понравиться
- Gentica CuantitativaДокумент28 страницGentica CuantitativaAlexis Sepulveda ValenzuelaОценок пока нет
- Genética Cuantitativa 2022Документ48 страницGenética Cuantitativa 2022Javier Arana100% (1)
- Practica N 08.-Genetica CuantitativaДокумент6 страницPractica N 08.-Genetica CuantitativaSelene CalleОценок пока нет
- Genetica CuantitativaДокумент67 страницGenetica CuantitativaElv ArévaloОценок пока нет
- Definiciones de Las Terminologias de Fitomejoramiento IДокумент3 страницыDefiniciones de Las Terminologias de Fitomejoramiento IAbimael Mao Sanchez VerdeОценок пока нет
- Pueraria Phseloides KudzuДокумент6 страницPueraria Phseloides KudzuLuis Fernando Schaus KochОценок пока нет
- Tipos de LarvasДокумент4 страницыTipos de LarvasJunior Montes ReynalteОценок пока нет
- Presentacion de Enf. PapilomaДокумент20 страницPresentacion de Enf. PapilomaFranklinCopaОценок пока нет
- Herencia Ligada Al SexoДокумент4 страницыHerencia Ligada Al Sexoydazp100% (1)
- Parametros ReproductivosДокумент4 страницыParametros ReproductivosOlman AlemanОценок пока нет
- Fitomejoramiento de Plantas AlógamasДокумент5 страницFitomejoramiento de Plantas Alógamasjohn100% (1)
- Informe 1 de GerminacionДокумент8 страницInforme 1 de GerminacionDavid FelixОценок пока нет
- King Grass Morado!Документ15 страницKing Grass Morado!Leydi CarreñoОценок пока нет
- Album Gramineas y LeguminosasДокумент50 страницAlbum Gramineas y LeguminosasAbdiel de Jesus Valencia HernandezОценок пока нет
- Diseños Bloques Incompletos2Документ31 страницаDiseños Bloques Incompletos2Pao Becerra E50% (2)
- King Grass MoradoДокумент7 страницKing Grass MoradoCarlos Alberto Fonseca BeltranОценок пока нет
- Microbiologia Del RumenДокумент20 страницMicrobiologia Del RumenDiego Armando Caupaz MartinesОценок пока нет
- Terminologia PasturasДокумент52 страницыTerminologia PasturasEstefania MoralesОценок пока нет
- Anatomía InsectilДокумент140 страницAnatomía InsectilFranklin J Urrutia Vidarte100% (1)
- Clase3 - Efecto de Un Solo GenДокумент35 страницClase3 - Efecto de Un Solo Genapi-3697648Оценок пока нет
- Poblemas de Genetica 3 Dominancia Incompleta Codominancia y Alelos LetalesДокумент3 страницыPoblemas de Genetica 3 Dominancia Incompleta Codominancia y Alelos LetalesAnonymous IHTo3Ijm100% (1)
- Vigor HibridoДокумент18 страницVigor Hibridofresh_2000Оценок пока нет
- UNIDAD 1 Genética MendelianaДокумент57 страницUNIDAD 1 Genética MendelianaSofia Guadalupe Lerma CarreónОценок пока нет
- Cromosomas de La PlantaДокумент2 страницыCromosomas de La PlantaBruno García GuillénОценок пока нет
- Pasto ElefanteДокумент26 страницPasto ElefanteEdy OmarОценок пока нет
- Deficiencias de Nutrientes en Las PlantasДокумент3 страницыDeficiencias de Nutrientes en Las PlantasGiorgio SánchezОценок пока нет
- Criterios para La Evaluación de PastizalesДокумент4 страницыCriterios para La Evaluación de PastizalesSantiagoGuerreroОценок пока нет
- Henolaje - Grupo 5Документ23 страницыHenolaje - Grupo 5Harold AgredaОценок пока нет
- Importancia de Los CuernosДокумент16 страницImportancia de Los CuernosLito Ram VerОценок пока нет
- Sesion 1 - PREPARACIÓN Y PARCELACIÓN DEL TERRENOДокумент11 страницSesion 1 - PREPARACIÓN Y PARCELACIÓN DEL TERRENOFátima Arias CabrejosОценок пока нет
- Determinacion Del Sexo 2Документ74 страницыDeterminacion Del Sexo 2Diustin Paredes Milla100% (2)
- La Importancia de La Meiosis en El Mejoramiento Genetico de Las PlantasДокумент4 страницыLa Importancia de La Meiosis en El Mejoramiento Genetico de Las PlantasAlonso Carrasco VásquezОценок пока нет
- Reconocimiento de Células SanguíneasДокумент8 страницReconocimiento de Células SanguíneasPaola Obando PadillaОценок пока нет
- Constantes Fisiológicas de Los Animales DomésticosДокумент2 страницыConstantes Fisiológicas de Los Animales Domésticositzel Guadalupe PereydaОценок пока нет
- Composicion de La Sangre de Animales Domesticos y Su FuncionДокумент2 страницыComposicion de La Sangre de Animales Domesticos y Su FuncionRaul Larios CurielОценок пока нет
- Problemas Mendelianos y No MendelianosДокумент5 страницProblemas Mendelianos y No MendelianosBertha Felix TarazonaОценок пока нет
- Ley de Hardy WeinbergДокумент6 страницLey de Hardy WeinbergLess NuñezОценок пока нет
- HeterosisДокумент9 страницHeterosisAustinОценок пока нет
- Tesis de Hibridacion MaizДокумент43 страницыTesis de Hibridacion MaizanthonyОценок пока нет
- 393276151.interacción Génica 2014 PDFДокумент20 страниц393276151.interacción Génica 2014 PDFMary Castillo100% (1)
- Crianza de Zabrotes Subfasciatus en Granos AlmacenadosДокумент9 страницCrianza de Zabrotes Subfasciatus en Granos AlmacenadosAstrid Távara VélezОценок пока нет
- Generalidades de La Reproducción en Ovinos y CaprinosДокумент57 страницGeneralidades de La Reproducción en Ovinos y CaprinosMarco Antonio López CarlosОценок пока нет
- Practica 1Документ3 страницыPractica 1Marybel Anahi Castillo FeratОценок пока нет
- Diseños Experimentales DCA - FactorialДокумент18 страницDiseños Experimentales DCA - FactorialJulio MalcaОценок пока нет
- Anatomía de La TruchaДокумент15 страницAnatomía de La TruchaRojas Jhon E100% (1)
- 08 Alelismo MultipleДокумент5 страниц08 Alelismo MultipleJorge Luis ParraОценок пока нет
- 1 La Propagacion Asexual NaturalДокумент33 страницы1 La Propagacion Asexual NaturalIvonne Elena Meza Bonett0% (1)
- Metabolismo Proteína RumiantesДокумент32 страницыMetabolismo Proteína RumiantesDesahily Juarez100% (1)
- Virus Del Mosaico Del PepinoДокумент4 страницыVirus Del Mosaico Del PepinoJessica Salcedo BerrioОценок пока нет
- MeloidogyneДокумент2 страницыMeloidogyneRonald RiveraОценок пока нет
- Tesis ForrajeДокумент131 страницаTesis ForrajeRicardo Suárez LópezОценок пока нет
- Transporte de AguaДокумент19 страницTransporte de AguapejeОценок пока нет
- Guia de Ejercicios Genética 2Документ4 страницыGuia de Ejercicios Genética 2Maribel AlvearОценок пока нет
- Diseños Con MEDIDAS REPETIDASДокумент19 страницDiseños Con MEDIDAS REPETIDASJennycitas EsquivelОценок пока нет
- 2 Genética Cualitativa, Defectos Genéticos PDFДокумент29 страниц2 Genética Cualitativa, Defectos Genéticos PDFBETTY JUDITH MACHACA ZAPANAОценок пока нет
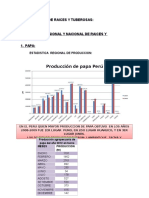
- Raices y TuberosasДокумент30 страницRaices y TuberosasVictor Peralta Quiroz100% (1)
- Polidipsia PoliuriaДокумент6 страницPolidipsia PoliuriaElianaFloresОценок пока нет
- Genetica Cuantitativa OkДокумент44 страницыGenetica Cuantitativa OkLuisMLОценок пока нет
- Genetica CuantitativaДокумент54 страницыGenetica CuantitativaAlexОценок пока нет
- Guia de Conceptos. Genetica Cuantitativa Ramirez - EgañaДокумент15 страницGuia de Conceptos. Genetica Cuantitativa Ramirez - EgañaElisabeth ReyesОценок пока нет
- Relatos-Guiado Museo 2016Документ28 страницRelatos-Guiado Museo 2016Danae TatajeОценок пока нет
- A7402-Evaluacion Hidrogeologica Ciudad Ilo-MoqueguaДокумент67 страницA7402-Evaluacion Hidrogeologica Ciudad Ilo-MoqueguaDanae TatajeОценок пока нет
- Descarga Constancia Fase Culminada-AdultosДокумент5 страницDescarga Constancia Fase Culminada-AdultosDanae TatajeОценок пока нет
- Bases de La Convocatoria - IcaДокумент6 страницBases de La Convocatoria - IcaDanae TatajeОценок пока нет
- Indicadores de Contaminación Del AguaДокумент19 страницIndicadores de Contaminación Del AguaDanae Tataje100% (1)
- Trámites para Optar El Título Profesional de Licenciado en La Especialidad Modalidad TesisДокумент2 страницыTrámites para Optar El Título Profesional de Licenciado en La Especialidad Modalidad TesisDanae TatajeОценок пока нет
- BIOPELÍCULASДокумент9 страницBIOPELÍCULASDanae TatajeОценок пока нет
- Informe de F. Vegetal N°1 Efecto de La Luz en La Germinación de Semillas de LechugaДокумент7 страницInforme de F. Vegetal N°1 Efecto de La Luz en La Germinación de Semillas de LechugaDanae TatajeОценок пока нет
- (Ok) Mycobacterium w2018Документ108 страниц(Ok) Mycobacterium w2018Danae TatajeОценок пока нет
- Capítulo 4 BiotecДокумент14 страницCapítulo 4 BiotecDanae TatajeОценок пока нет
- Bio SeguridadДокумент5 страницBio SeguridadDanae TatajeОценок пока нет
- Guia de Usuario Descargable PDFДокумент24 страницыGuia de Usuario Descargable PDFKenyo Loa100% (1)
- Capítulo 5 BiotecДокумент11 страницCapítulo 5 BiotecDanae TatajeОценок пока нет
- Normas LegalesДокумент14 страницNormas LegalesDanae TatajeОценок пока нет
- Original Article / Artículo OriginalДокумент17 страницOriginal Article / Artículo OriginalDanae TatajeОценок пока нет
- Hongos Encontrados en Árboles Del Campus de La UnfvДокумент20 страницHongos Encontrados en Árboles Del Campus de La UnfvDanae TatajeОценок пока нет
- Actual Norma Criterios Microbiologicos Alimentos RM591MINSAДокумент26 страницActual Norma Criterios Microbiologicos Alimentos RM591MINSAedmilos100% (4)
- Enfermedad de HuntingtonДокумент4 страницыEnfermedad de HuntingtonDanae TatajeОценок пока нет
- En CuestaДокумент1 страницаEn CuestaDanae TatajeОценок пока нет
- Bioseguridad NivelesДокумент6 страницBioseguridad NivelesLuis Enrique AlvaradoОценок пока нет
- LibroRojo Ballenajorobada 2016-07-25Документ5 страницLibroRojo Ballenajorobada 2016-07-25Danae TatajeОценок пока нет
- 1 Historia de La MicrobiologiaДокумент33 страницы1 Historia de La MicrobiologiafionamariposaОценок пока нет
- 1 Historia de La MicrobiologiaДокумент33 страницы1 Historia de La MicrobiologiafionamariposaОценок пока нет
- 0 - Historia Microbiologia 2013Документ108 страниц0 - Historia Microbiologia 2013Danae TatajeОценок пока нет
- Sem 101Документ181 страницаSem 101Miguel RivasОценок пока нет
- cPITULO 7, PERSONALIDADДокумент38 страницcPITULO 7, PERSONALIDADMaria Luengo DiazОценок пока нет
- 09-Catalogos de Bovinos LecherosДокумент14 страниц09-Catalogos de Bovinos LecherosDany Sagrero García0% (1)
- Herencia y Heredabilidad A Y CДокумент11 страницHerencia y Heredabilidad A Y CItzelОценок пока нет
- La Heredabilidad en Las Ciencias Del DeporteДокумент7 страницLa Heredabilidad en Las Ciencias Del DeporteCristian VásquezОценок пока нет
- Psicología Evolutiva Tema 1Документ5 страницPsicología Evolutiva Tema 1Jota Jota TrilobiteОценок пока нет
- Índices de Heredabilidad (Genética)Документ2 страницыÍndices de Heredabilidad (Genética)marta madridОценок пока нет
- Genética de La ConductaДокумент3 страницыGenética de La ConductaEcbОценок пока нет
- Heredabilidad PDFДокумент9 страницHeredabilidad PDFErika FC100% (3)
- Genetica Aplicada A La Produccion de Bovinos de CarneДокумент11 страницGenetica Aplicada A La Produccion de Bovinos de CarneCZCASEZATОценок пока нет
- Apuntes de La AsignaturaДокумент76 страницApuntes de La AsignaturaMer AriasОценок пока нет
- Mejoramiento GeneticoДокумент24 страницыMejoramiento GeneticoBreiner Andrés Trujillo PérezОценок пока нет
- Reproducción Selectiva de Especies Forestales de Rapido CrecimientoДокумент68 страницReproducción Selectiva de Especies Forestales de Rapido CrecimientoMarcelo Hernandez Nauto0% (1)
- Consejería Genética - López Diego y Méndez Steven - PreterminadoДокумент55 страницConsejería Genética - López Diego y Méndez Steven - PreterminadoSTEVEN WILTER JUNIOR MENDEZ ALVARADOОценок пока нет
- Fitomejoramiento - Herencia Cualitativa y Cuantitativa, Unidad Ii, Semana 9Документ25 страницFitomejoramiento - Herencia Cualitativa y Cuantitativa, Unidad Ii, Semana 9Euler Jhonatan Córdova RojasОценок пока нет
- Etapa 7 - Prueba Final, FITOMEJORAMIENTO PDFДокумент15 страницEtapa 7 - Prueba Final, FITOMEJORAMIENTO PDFAlexis JanamanchichoyОценок пока нет
- Bos Taurus y Bos Indicus Mejoramiento GeneticoДокумент30 страницBos Taurus y Bos Indicus Mejoramiento GeneticoGabriel ViteОценок пока нет
- Capitulo Vii. SeleccionДокумент49 страницCapitulo Vii. SeleccionElmer Meza RojasОценок пока нет
- El Agora de La NeuroeducaciónДокумент260 страницEl Agora de La NeuroeducaciónMario Bross92% (12)
- Guia de Ejercicios Practico 8Документ4 страницыGuia de Ejercicios Practico 8Lipu BolliniОценок пока нет
- OvinosДокумент105 страницOvinosManuhg HgОценок пока нет
- Tema 3. Influencias Genéticas y Culturales en PersonalidadДокумент32 страницыTema 3. Influencias Genéticas y Culturales en PersonalidadandreaОценок пока нет
- Preguntas y Respuestas Sobre Las DEPs en AlpacasДокумент6 страницPreguntas y Respuestas Sobre Las DEPs en AlpacasArmando Fernandez Luna100% (1)
- Fitomejoramiento - Heredabilidad y Ligamiento Factorial, Unidad Ii, Semana 9, Ciclo 2020-Ii BДокумент14 страницFitomejoramiento - Heredabilidad y Ligamiento Factorial, Unidad Ii, Semana 9, Ciclo 2020-Ii Bdelvis sanchez diasОценок пока нет
- Etapa 2 Bases FitomejoramientoДокумент10 страницEtapa 2 Bases FitomejoramientoJaider eduardo Osorio ragoОценок пока нет
- BQCGFHДокумент317 страницBQCGFHElena Palomo CastroОценок пока нет
- PRÁCTICA 3 - PROBLEMAS DE HEREDABILIDAD Mejoramiento GeneticoДокумент4 страницыPRÁCTICA 3 - PROBLEMAS DE HEREDABILIDAD Mejoramiento Geneticomarco maldonadoОценок пока нет
- Heredabilidad de Las Caracteristicas Psicologicas OkДокумент9 страницHeredabilidad de Las Caracteristicas Psicologicas OkJohana PatiñoОценок пока нет
- Bases Teóricas para El Mejoramiento Genético de TilapiasДокумент110 страницBases Teóricas para El Mejoramiento Genético de TilapiasTania Cristina Muñoz Nuñez100% (1)
- 250 Guzera EspanholДокумент102 страницы250 Guzera EspanholDiana Catalina Castiblanco SorianoОценок пока нет
- Cuestionario RespuestasДокумент10 страницCuestionario RespuestasEdinson Padilla Pajaro100% (2)