Вам также может понравиться
- Complejo Mayor de Histocompatibilidad JuanДокумент27 страницComplejo Mayor de Histocompatibilidad JuanJuan Sarid Domínguez OrtízОценок пока нет
- Complejo Mayor de Histocompatibilidad 2Документ58 страницComplejo Mayor de Histocompatibilidad 2Raquel Tejada LantiguaОценок пока нет
- HLAДокумент24 страницыHLAGisselle Soto MinchaloОценок пока нет
- Complejomayordehistocompatibilidad 1 120908001144 Phpapp01Документ53 страницыComplejomayordehistocompatibilidad 1 120908001144 Phpapp01Voltaire Méndez RodríguezОценок пока нет
- Complejo Mayor de HistocompatibilidadДокумент6 страницComplejo Mayor de HistocompatibilidadRaquel GuevaraОценок пока нет
- I Hla PostДокумент51 страницаI Hla PostDavid Fernando Cujuy SuarezОценок пока нет
- 8 - Complejo Mayor de HistocompatibildiadДокумент11 страниц8 - Complejo Mayor de HistocompatibildiadOzxiliusОценок пока нет
- HLA InmunologiaДокумент29 страницHLA InmunologiaJunior AlencarОценок пока нет
- Sistema InmuneДокумент16 страницSistema InmuneRaquel EspinozaОценок пока нет
- Complejo Mayor de HistocompatibilidadДокумент20 страницComplejo Mayor de HistocompatibilidadDaniel HerreraОценок пока нет
- InmonologiaДокумент36 страницInmonologiaOsvaldo SantiagoОценок пока нет
- APC SeminarioДокумент24 страницыAPC SeminarioAnne NikolОценок пока нет
- Artículo MHCДокумент3 страницыArtículo MHCLuis TorresОценок пока нет
- Tema 5 Complejo Mayor HistocompatibilidadДокумент19 страницTema 5 Complejo Mayor HistocompatibilidadANGEL BAQUEОценок пока нет
- Hla 2017 InmunoДокумент36 страницHla 2017 InmunoMonica OrdinolaОценок пока нет
- Sistema Mayor de HistocompatibilidadДокумент28 страницSistema Mayor de HistocompatibilidadCarlosJuniorCalleОценок пока нет
- Complejo MHДокумент33 страницыComplejo MHNasha Ardila'Оценок пока нет
- Presentacion de Antigenos - Pre. 3Документ13 страницPresentacion de Antigenos - Pre. 3Angelica Raquel Diaz Corrales0% (1)
- Complejo de HistocompatibilidadДокумент1 страницаComplejo de HistocompatibilidadRUTH ANGELICA BAEZ JARAMILLOОценок пока нет
- Resumen Inmunologia Introduccion y GeneralidadesДокумент15 страницResumen Inmunologia Introduccion y GeneralidadesJoel SánchezОценок пока нет
- Seminario III - InmunoДокумент7 страницSeminario III - InmunoAlexander Chappa MaiceloОценок пока нет
- Capitulo 3 Abbas Captura Del Antigeno y Presentacion A Los Linfocitos 1 DownloableДокумент5 страницCapitulo 3 Abbas Captura Del Antigeno y Presentacion A Los Linfocitos 1 DownloableSergio Fernando Cruz100% (1)
- Articulo MCHДокумент7 страницArticulo MCHNathaly DuarteОценок пока нет
- Receptores de Reconocimiento Celular Antigenico. InmunologiaДокумент13 страницReceptores de Reconocimiento Celular Antigenico. Inmunologiaadalbertop50% (2)
- Libro DR Huanqui - CompressДокумент141 страницаLibro DR Huanqui - CompressValery Mariana Olivera ZunigaОценок пока нет
- 8 MHC 2020Документ33 страницы8 MHC 2020Luna GabrielaОценок пока нет
- MHCДокумент29 страницMHCscribd_slideworksОценок пока нет
- Revisión Bibliográfica de Complejo Mayor de HistocompatibilidadДокумент5 страницRevisión Bibliográfica de Complejo Mayor de HistocompatibilidadOtoney Intriago100% (2)
- Complejo Mayor de Histocompatibilidad. RevistaFacMed09Документ4 страницыComplejo Mayor de Histocompatibilidad. RevistaFacMed09hadita31vmОценок пока нет
- Cap 3. Captura Del Antígeno y Presentación A Los LinfocitosДокумент8 страницCap 3. Captura Del Antígeno y Presentación A Los LinfocitosSebastián FranciscoОценок пока нет
- Complejo de HistocompatibilidadДокумент25 страницComplejo de HistocompatibilidadVALERIA YARENTZI CASTRO MOROОценок пока нет
- Antígenos de HistocompatibilidadДокумент29 страницAntígenos de HistocompatibilidadEinar Eduardo Fuentes ZamudioОценок пока нет
- Complejo Mayor de Histocompatibilidad Julio Ruiz Quiroz 2012Документ39 страницComplejo Mayor de Histocompatibilidad Julio Ruiz Quiroz 2012lucio obandoОценок пока нет
- Trabajo Final de CPH 1Документ19 страницTrabajo Final de CPH 1Selene Vanessa Chozo Mestanza0% (1)
- CHPДокумент34 страницыCHPCARNAGEОценок пока нет
- Taller de Biofísica 2 Complejo Mayor de HistocompatibilidadДокумент8 страницTaller de Biofísica 2 Complejo Mayor de HistocompatibilidadAlejandra VelasquezОценок пока нет
- COMPLEJO MAYOR DE HISTOCOMPATIBILIDAD y TIPIFICACION DE HLAДокумент13 страницCOMPLEJO MAYOR DE HISTOCOMPATIBILIDAD y TIPIFICACION DE HLASebastian aguilar RodríguezОценок пока нет
- Complejo Mayor de HistocompatibilidadДокумент40 страницComplejo Mayor de HistocompatibilidadLeida Garcia RamirezОценок пока нет
- Presentacion AntigenicaДокумент130 страницPresentacion AntigenicaCarlaОценок пока нет
- Apuntes Inmunologia: Primer Parcial: Tema 1: IntroducciónДокумент38 страницApuntes Inmunologia: Primer Parcial: Tema 1: IntroducciónAlexshaguОценок пока нет
- Libro Inmuno HUANQUIДокумент140 страницLibro Inmuno HUANQUIAngelicaОценок пока нет
- Sem 5 InmunДокумент4 страницыSem 5 InmunVictoria CortésОценок пока нет
- Complejo Mayor de Histocompatibilidad (MHC)Документ6 страницComplejo Mayor de Histocompatibilidad (MHC)Laura TorresОценок пока нет
- Complejo Mayor de HistocompatibilidadДокумент15 страницComplejo Mayor de HistocompatibilidadAndrea RangelОценок пока нет
- 1.4 CÉLULA PRESENTADORA DE ANTÍGENOS ModДокумент70 страниц1.4 CÉLULA PRESENTADORA DE ANTÍGENOS ModBy Fran JoséОценок пока нет
- Capitulo 9 InmunoДокумент18 страницCapitulo 9 InmunoYarlen MoralesОценок пока нет
- Presentacion de AntígenosДокумент8 страницPresentacion de AntígenosJosé Manuel mojica villamizarОценок пока нет
- MHC GeneralidadesДокумент9 страницMHC GeneralidadesAldo AguilarОценок пока нет
- Taller CMHДокумент3 страницыTaller CMHLaura Sofia Velandia Naranjo100% (1)
- Resumen MHCДокумент3 страницыResumen MHCLuis GuzmánОценок пока нет
- Resumen Capitulo Inmunología RobbinsДокумент17 страницResumen Capitulo Inmunología RobbinsLaura Galvis33% (3)
- Celulas Presentadoras de Anrigenos (Apc) : Células Presentadoras ProfesionalesДокумент14 страницCelulas Presentadoras de Anrigenos (Apc) : Células Presentadoras ProfesionaleslipgarОценок пока нет
- Complejo CMHДокумент7 страницComplejo CMHandrea.med2401Оценок пока нет
- Inmunidad AdaptativaДокумент4 страницыInmunidad AdaptativaGiudiche BrisaОценок пока нет
- Complejo Mayor de HistocompatibilidadДокумент47 страницComplejo Mayor de Histocompatibilidadnomade.astralОценок пока нет
- Complejo Mayor de Histocompatibilidad - ResumenДокумент1 страницаComplejo Mayor de Histocompatibilidad - ResumenMagda AguilarОценок пока нет
- Investiga RДокумент5 страницInvestiga RCarlos Eduardo Díaz MoralesОценок пока нет
- Introducción a la antropología biológica: Origen, variabilidad y adaptación de las poblaciones humanasОт EverandIntroducción a la antropología biológica: Origen, variabilidad y adaptación de las poblaciones humanasОценок пока нет
- Fortalecer El Tibial AnteriorДокумент4 страницыFortalecer El Tibial AnteriorErwin Hernández García100% (1)
- Diferencias Entre Tanque de Whirlpool y HubbardДокумент10 страницDiferencias Entre Tanque de Whirlpool y HubbardErwin Hernández García100% (1)
- Introduccion Cinesiterapia - HemiplejìaДокумент14 страницIntroduccion Cinesiterapia - HemiplejìaErwin Hernández GarcíaОценок пока нет
- Biotecnología en La Producción de AntibióticosДокумент8 страницBiotecnología en La Producción de AntibióticosErwin Hernández GarcíaОценок пока нет
- Exposicion de Arte Danzas Tipicas de La SierraДокумент16 страницExposicion de Arte Danzas Tipicas de La SierraErwin Hernández GarcíaОценок пока нет
- CAUSAS LumbalgiaДокумент2 страницыCAUSAS LumbalgiaErwin Hernández GarcíaОценок пока нет
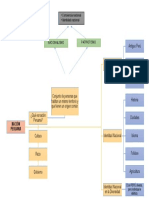
- Mapa Conceptual Nación PeruanaДокумент1 страницаMapa Conceptual Nación PeruanaErwin Hernández García67% (3)
- El MuñónДокумент3 страницыEl MuñónErwin Hernández GarcíaОценок пока нет
- Encuesta de Las 3R y Los Residuos SólidosДокумент1 страницаEncuesta de Las 3R y Los Residuos SólidosErwin Hernández García100% (1)
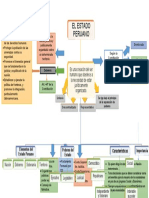
- Mapa Conceptual Estado PeruanoДокумент1 страницаMapa Conceptual Estado PeruanoErwin Hernández García100% (4)
- Mapa Conceptual La SociedadДокумент1 страницаMapa Conceptual La SociedadErwin Hernández García0% (1)
- Signos VitalesДокумент31 страницаSignos VitalesErwin Hernández GarcíaОценок пока нет
- HuaycoloroДокумент2 страницыHuaycoloroErwin Hernández GarcíaОценок пока нет
- Barras Paralelas para RehabilitacionДокумент9 страницBarras Paralelas para RehabilitacionErwin Hernández GarcíaОценок пока нет
- Practica AINESДокумент5 страницPractica AINESErwin Hernández GarcíaОценок пока нет
- 11 MecanoterapiaДокумент18 страниц11 MecanoterapiaErwin Hernández GarcíaОценок пока нет
- Virus de Plantas ExposiciónДокумент47 страницVirus de Plantas ExposiciónErwin Hernández García0% (1)
- InmunosuerosДокумент23 страницыInmunosuerosErwin Hernández GarcíaОценок пока нет
- Diseños y Bioprocesos de Los AntibioticosДокумент12 страницDiseños y Bioprocesos de Los AntibioticosErwin Hernández GarcíaОценок пока нет
- Ciclo de Replicación ViralДокумент41 страницаCiclo de Replicación ViralJohan VásquezОценок пока нет
- Clases 15-16 Ciclo Celular y MitosisДокумент93 страницыClases 15-16 Ciclo Celular y MitosisKAREN MONSERRAT FRANCO ACHARОценок пока нет
- Tarea V Biologia y Su Lab... Emei BelloДокумент32 страницыTarea V Biologia y Su Lab... Emei BelloESMARLIS BELLO JOSE100% (1)
- La REPRODUCCIÓN ASEXUAL y SEXUALДокумент2 страницыLa REPRODUCCIÓN ASEXUAL y SEXUALByron ZumОценок пока нет
- SodapdfДокумент1 страницаSodapdfCamilo HerreraОценок пока нет
- Componentes ProtoplasmaticosДокумент8 страницComponentes ProtoplasmaticosLuis Gustavo Guillen100% (1)
- 1.introducción Al Sistema InmuneДокумент6 страниц1.introducción Al Sistema InmuneIsabella CastilloОценок пока нет
- Guía de Lectura Potenciales de MembranaДокумент3 страницыGuía de Lectura Potenciales de Membranaisabellaprie3Оценок пока нет
- Actividades - Biologia para Imprimir PDFДокумент18 страницActividades - Biologia para Imprimir PDFJosafatMasterMartinezMartinezОценок пока нет
- Dogma Central de La Biología MolecularДокумент3 страницыDogma Central de La Biología MolecularGUZMAN HERNANDEZ MARIA GUADALUPEОценок пока нет
- El Reino Protista Protozoos para Segundo de SecundariaДокумент8 страницEl Reino Protista Protozoos para Segundo de SecundariaDanhia Velarde HuinchoОценок пока нет
- 11 - Fisiologia LeucocitosДокумент53 страницы11 - Fisiologia Leucocitosmariajose100% (1)
- EDA - 06 - 4º - ACTIVIDAD 01 - CyT - 2022Документ14 страницEDA - 06 - 4º - ACTIVIDAD 01 - CyT - 2022Juan Acostupa CahuanaОценок пока нет
- LipidosДокумент2 страницыLipidosSaraОценок пока нет
- Metabolismo de Acilgliceroles y EsfingolípidosДокумент36 страницMetabolismo de Acilgliceroles y EsfingolípidosSebastián RivadeneiraОценок пока нет
- Inhibición EnzimáticaДокумент31 страницаInhibición EnzimáticaKevin SalazarОценок пока нет
- Examen Inmunologia IIIДокумент7 страницExamen Inmunologia IIILidia Romero ValeroОценок пока нет
- Inflamación Aguda 2020 PDFДокумент34 страницыInflamación Aguda 2020 PDFAndrea Garcia CaycedoОценок пока нет
- Ing. EnzimáticaДокумент46 страницIng. EnzimáticaArmando Montoya García100% (1)
- Examen Tipo ICFES Biología 11Документ2 страницыExamen Tipo ICFES Biología 11JhonEspejoBayonaОценок пока нет
- Impulso NerviosoДокумент1 страницаImpulso NerviosoDyker Evaj100% (4)
- Taller BiologíaДокумент4 страницыTaller Biologíanaruto10170% (1)
- Informe Practica de Microscopia Extendido de SangreДокумент4 страницыInforme Practica de Microscopia Extendido de SangreValentina RestrepoОценок пока нет
- Grupo 11 Tejido Hematopoyético PreguntasДокумент2 страницыGrupo 11 Tejido Hematopoyético PreguntasMateoo06Оценок пока нет
- Diferentes Estructuras de Las Proteinas 2Документ2 страницыDiferentes Estructuras de Las Proteinas 2Pedro Javier Barrios OrmeñoОценок пока нет
- Resumen ParkinsonДокумент6 страницResumen ParkinsonLuissita CriolloОценок пока нет
- 3 Membrana y TransporteДокумент32 страницы3 Membrana y TransporteMaria Jose ROОценок пока нет
- Mus CuloДокумент34 страницыMus CuloAbel GonzalezОценок пока нет
- Investigación de Las Células Eucariotas y AnimalesДокумент5 страницInvestigación de Las Células Eucariotas y AnimalesJorge Luis Monge MendozaОценок пока нет
- ARLETHДокумент2 страницыARLETHArleth SalazarОценок пока нет