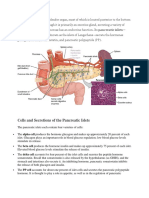
Вам также может понравиться
- CHAPTER 22 Gluconeogenesis, Glycogen Metabolism, and The Pentose Phosphate PathwayДокумент12 страницCHAPTER 22 Gluconeogenesis, Glycogen Metabolism, and The Pentose Phosphate Pathway楊畯凱Оценок пока нет
- Gluconeogenesis and Cori CycleДокумент21 страницаGluconeogenesis and Cori CycleAboubakar Moalim Mahad moh'dОценок пока нет
- Hexose Mono Phosphate (HMP) ShuntДокумент28 страницHexose Mono Phosphate (HMP) ShuntVishesh JainОценок пока нет
- Gluconeogenesis Metabolic PathwayДокумент36 страницGluconeogenesis Metabolic PathwayDayledaniel Sorveto0% (1)
- 13 - GluconeogenesisДокумент23 страницы13 - GluconeogenesischeckmateОценок пока нет
- Integration of Metabolism PathwaysДокумент68 страницIntegration of Metabolism PathwaysCahyani Tiara Safitri100% (1)
- Metabolism of Purine & Pyrimidine NucleotidesДокумент38 страницMetabolism of Purine & Pyrimidine NucleotidesShimmering MoonОценок пока нет
- Metabolic Pathway of Carbohydrate and GlycolysisДокумент22 страницыMetabolic Pathway of Carbohydrate and GlycolysisDarshansinh MahidaОценок пока нет
- GlycogenolysisДокумент37 страницGlycogenolysisJasmine Rey QuintoОценок пока нет
- Digestion and Absorption of Dietary LipidsДокумент35 страницDigestion and Absorption of Dietary LipidsCharmaine Luc100% (2)
- Digestion and Absorption of Carbohydrates: by Arpitha 3 BnysДокумент31 страницаDigestion and Absorption of Carbohydrates: by Arpitha 3 BnysBig ZeroОценок пока нет
- Understanding Metabolism of CarbohydratesДокумент48 страницUnderstanding Metabolism of CarbohydratesAbdullah TheОценок пока нет
- Glycolysis NotesДокумент2 страницыGlycolysis NotesAnsley Ellen BarfieldОценок пока нет
- Fatty Acid Synthesis 11.12.19Документ18 страницFatty Acid Synthesis 11.12.19Sanreet Randhawa100% (1)
- Hexose Monophosphate Shunt CHEM3119"TITLE"Biometabolism Lecture on HMP Shunt and G6PD DeficiencyДокумент18 страницHexose Monophosphate Shunt CHEM3119"TITLE"Biometabolism Lecture on HMP Shunt and G6PD DeficiencyAbdul Jabbar Abdul JabbarОценок пока нет
- Clinical SignificanceДокумент31 страницаClinical Significancejav israelОценок пока нет
- Lipoprotein MetabolismДокумент23 страницыLipoprotein MetabolismDarien LiewОценок пока нет
- Glucose Homeostasis and DiabetesДокумент33 страницыGlucose Homeostasis and DiabetesSarahОценок пока нет
- Pancreatic Islets Hormones Regulate Blood GlucoseДокумент4 страницыPancreatic Islets Hormones Regulate Blood GlucoseSophia OcayОценок пока нет
- Gluconeogenesis & HMP Shunt ExplainedДокумент36 страницGluconeogenesis & HMP Shunt ExplainedAshish K Joy100% (1)
- Glycogen MetabolismДокумент11 страницGlycogen MetabolismSimra ZahidОценок пока нет
- Biochem HomeworkДокумент13 страницBiochem Homeworkfcukingfranztastik50% (2)
- Amino Acid Catabolism Between OrgansДокумент44 страницыAmino Acid Catabolism Between OrgansFarhati MardhiyahОценок пока нет
- Chapter 24 Practice QuestionsДокумент7 страницChapter 24 Practice QuestionsArlene F. Montalbo100% (1)
- Carbohydrates SummaryДокумент9 страницCarbohydrates SummaryHarold NagunaОценок пока нет
- HW - Carbohydrate Metabolism II & Lipid MetabolismДокумент2 страницыHW - Carbohydrate Metabolism II & Lipid MetabolismyanОценок пока нет
- CHAPTER 18 GlycolysisДокумент10 страницCHAPTER 18 Glycolysis楊畯凱Оценок пока нет
- Lipids LehningerДокумент7 страницLipids LehningerElla BangalanОценок пока нет
- 8 Carbohydrate Metabolism PDFДокумент36 страниц8 Carbohydrate Metabolism PDFDayne Ocampo-SolimanОценок пока нет
- Some Answer of Problemset - 7 - KEYДокумент3 страницыSome Answer of Problemset - 7 - KEYNihir PatelОценок пока нет
- Biochemistry-Exam 2 Heinz SchwarzkopfДокумент162 страницыBiochemistry-Exam 2 Heinz SchwarzkopfthomasОценок пока нет
- Biological Oxidation Pathways and EnzymesДокумент46 страницBiological Oxidation Pathways and EnzymesTelesha Zabie50% (2)
- GlycogenesisДокумент22 страницыGlycogenesisDaniel IvanОценок пока нет
- Ketone BodiesДокумент16 страницKetone BodiesAsim Ali100% (1)
- HMPДокумент44 страницыHMPraanja2Оценок пока нет
- Also Known As TCA Cycle Essentially Involves This Is The Final Common Oxidative Pathway ForДокумент10 страницAlso Known As TCA Cycle Essentially Involves This Is The Final Common Oxidative Pathway ForNarayan debnath hridoyОценок пока нет
- Amino SugarsДокумент2 страницыAmino SugarsNarasimha MurthyОценок пока нет
- Practical BiochemistryДокумент35 страницPractical BiochemistryMockinjay100% (1)
- Fa Lipid MetabДокумент4 страницыFa Lipid MetabJoy VergaraОценок пока нет
- Glycolysis: Shekhar Chandra Yadav Lecturer Dept. of BiochemistryДокумент25 страницGlycolysis: Shekhar Chandra Yadav Lecturer Dept. of BiochemistryTULSI SHARMAОценок пока нет
- S13.CHO .MetabolismДокумент18 страницS13.CHO .MetabolismGhea Jovita SinagaОценок пока нет
- Phenylalanine and Tyrosine Metabolism (18 Oct)Документ14 страницPhenylalanine and Tyrosine Metabolism (18 Oct)api-19824406100% (6)
- Beta OxidationДокумент19 страницBeta Oxidationindra100% (1)
- MCQ MembranesДокумент10 страницMCQ MembranesMarilyne RizkОценок пока нет
- (مكتملة) اسئلة البايو د.ج السمهري (.)Документ10 страниц(مكتملة) اسئلة البايو د.ج السمهري (.)Ozgan SüleymanОценок пока нет
- SummaryДокумент21 страницаSummarydindaОценок пока нет
- Insulin and Glucagon Regulation of Metabolic HomeostasisДокумент40 страницInsulin and Glucagon Regulation of Metabolic HomeostasisEka HabinaОценок пока нет
- Urea Cycle: DR Fasiha Fatima Department of BiochemistryДокумент30 страницUrea Cycle: DR Fasiha Fatima Department of BiochemistryAmmar YasirОценок пока нет
- FAT Soluble VitaminsДокумент53 страницыFAT Soluble VitaminsFaria LatifОценок пока нет
- Metabolism of Carbohydrate: Department of Biochemistry Faculty of Medicine University of YARSI JakartaДокумент60 страницMetabolism of Carbohydrate: Department of Biochemistry Faculty of Medicine University of YARSI JakartaAmanda PutriОценок пока нет
- Carbohydrate MetabolismДокумент57 страницCarbohydrate MetabolismYulliza Kurniawaty L100% (1)
- Heme Metabolism PDFДокумент19 страницHeme Metabolism PDFAnonymous jW7BU44ACОценок пока нет
- Carbohydrate Metabolism: Blood Haemoglobin and Respiration: - Archit SaraogiДокумент6 страницCarbohydrate Metabolism: Blood Haemoglobin and Respiration: - Archit SaraogiChictoran RukmaniОценок пока нет
- Biochemistry TestДокумент2 страницыBiochemistry Testbrijeshandfamily100% (1)
- Glycoproteins and Proteoglycans TTVДокумент34 страницыGlycoproteins and Proteoglycans TTVcraigОценок пока нет
- Metabolism of DisaccharidesДокумент15 страницMetabolism of DisaccharidesminaОценок пока нет
- 4 Bioenergetics and Oxidative Metabolism IIДокумент3 страницы4 Bioenergetics and Oxidative Metabolism IILinus LiuОценок пока нет
- Beta Oxidation of Fatty AcidsДокумент34 страницыBeta Oxidation of Fatty AcidsZelda LoverОценок пока нет
- Gluconeogenesis Pathway and Significance in 40 CharactersДокумент31 страницаGluconeogenesis Pathway and Significance in 40 CharactersTahera AhmedОценок пока нет
- Micronutrients in health and disease reviewДокумент10 страницMicronutrients in health and disease reviewLeonard LeonardОценок пока нет
- Glycolysis and TCA CycleДокумент38 страницGlycolysis and TCA CycleAboubakar Moalim Mahad moh'dОценок пока нет
- Unit 6 WorksheetДокумент6 страницUnit 6 WorksheetMarta YantiОценок пока нет
- Beta OxidationДокумент10 страницBeta OxidationAiane Gramatica100% (1)
- Metabolism of Purine & Pyrimidine NucleotidesДокумент38 страницMetabolism of Purine & Pyrimidine NucleotidesShimmering MoonОценок пока нет
- Biology Class 9th NotesДокумент20 страницBiology Class 9th NotesShakeel Akhtar AwanОценок пока нет
- Metabolic Acidosis FinalДокумент29 страницMetabolic Acidosis FinalTera SurbaktiОценок пока нет
- CH407 Biochemical EngineeringДокумент2 страницыCH407 Biochemical EngineeringVish VishwasОценок пока нет
- Enzymatic and Non EnzymaticДокумент28 страницEnzymatic and Non EnzymaticHaseen AhmedОценок пока нет
- Energy For All!: Engage: Food For CornДокумент4 страницыEnergy For All!: Engage: Food For CornBri LavaeОценок пока нет
- Nutrient Chart Deficiency and ToxicityДокумент7 страницNutrient Chart Deficiency and Toxicityapi-256577647Оценок пока нет
- Nutrient Timing For Peak PerformanceДокумент248 страницNutrient Timing For Peak Performancemd91101100% (2)
- Nature's Silent HealerДокумент10 страницNature's Silent HealerEager BeaverОценок пока нет
- Ehninger Principles of Biochemistry, Fourth Edition - DavidNelson, Michael M. Cox (0641-0680) PDFДокумент40 страницEhninger Principles of Biochemistry, Fourth Edition - DavidNelson, Michael M. Cox (0641-0680) PDFDesmon Jonathan SumolangОценок пока нет
- ETC Electron Transport Chain Lab Manual (English)Документ8 страницETC Electron Transport Chain Lab Manual (English)jakeyy harukyuОценок пока нет
- 2.1 Molecules To MetabolismДокумент44 страницы2.1 Molecules To MetabolismJianing Claire LiОценок пока нет
- Protein MetabolismДокумент15 страницProtein MetabolismAfshan Akhtar100% (1)
- INTRODUCTION TO HUMAN NUTRITIONДокумент35 страницINTRODUCTION TO HUMAN NUTRITIONNANCYОценок пока нет
- NORD Physician Guide To Urea Cycle DisordersДокумент28 страницNORD Physician Guide To Urea Cycle DisordersUzair TariqОценок пока нет
- Amino Acid Metabolism and the Urea CycleДокумент3 страницыAmino Acid Metabolism and the Urea Cyclet-oОценок пока нет
- Nutrient Analysis - Lab Report - BiochemДокумент3 страницыNutrient Analysis - Lab Report - BiochemdzdooОценок пока нет
- The Nitrogen Cycle: Processes, Players, and Human ImpactДокумент8 страницThe Nitrogen Cycle: Processes, Players, and Human ImpactKesto LoriakОценок пока нет
- Na Mlbio Ch10Документ28 страницNa Mlbio Ch10itaemin243Оценок пока нет
- Bgm, A Newly Synthesized Boron Compound, Induces Apoptosis And Reduces Oxidative Stress By Inhibiting Lipogenesis In 3T3-L1 Adipocytes Via Pparγ And Ctrp3Документ20 страницBgm, A Newly Synthesized Boron Compound, Induces Apoptosis And Reduces Oxidative Stress By Inhibiting Lipogenesis In 3T3-L1 Adipocytes Via Pparγ And Ctrp3eric smidtОценок пока нет
- Natural Anti Viral Self Defense StevenWmFowkesДокумент251 страницаNatural Anti Viral Self Defense StevenWmFowkesChristopher Phillips100% (1)
- Lecture Presentation - Protein MetabolismДокумент8 страницLecture Presentation - Protein Metabolismmr.avdheshsharma100% (1)
- Feeder Pathways For GlycolysisДокумент62 страницыFeeder Pathways For GlycolysisAbdikafar AbdullahiОценок пока нет
- Research: Lanosterol to Cholesterol PathwayДокумент8 страницResearch: Lanosterol to Cholesterol PathwaymonicaОценок пока нет
- Improvement of Shelf Life of Injera' Using Chemical PreservativesДокумент86 страницImprovement of Shelf Life of Injera' Using Chemical Preservativesfikeryk100% (4)
- Bioestimulantes - Generalidades, Definiciones, Mecanismos.Документ32 страницыBioestimulantes - Generalidades, Definiciones, Mecanismos.Jocelyn BelenoОценок пока нет