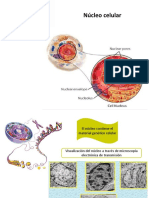
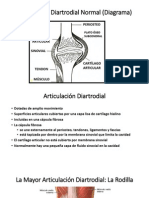
Вам также может понравиться
- Guia de Practicas - Genetica y Mejoramiento Animal - Carlos Scotto Espinoza - 2019Документ68 страницGuia de Practicas - Genetica y Mejoramiento Animal - Carlos Scotto Espinoza - 2019Claudia Castillo IngarОценок пока нет
- Prueba. III-IV° Año Medio. Plan Diferenciado. 2022. Ciclo Celular, Replicación, Transcripción y Traducción. Forma A. SCДокумент12 страницPrueba. III-IV° Año Medio. Plan Diferenciado. 2022. Ciclo Celular, Replicación, Transcripción y Traducción. Forma A. SCMauricio OrellanaОценок пока нет
- Los VirusДокумент3 страницыLos Virusdanii22180% (1)
- Clase Estructura Del NucleoДокумент90 страницClase Estructura Del Nucleohghbkgykmgxyh100% (1)
- Nucleo CelularДокумент34 страницыNucleo CelularPedro Durand EspinozaОценок пока нет
- Conferencia Biologia Nivelación Núcleo-1Документ25 страницConferencia Biologia Nivelación Núcleo-1Dome BenavidesОценок пока нет
- NucleoДокумент39 страницNucleoEdwin QuizhpiОценок пока нет
- Núcleo Celular 2021Документ47 страницNúcleo Celular 2021María Nathalie Hidálgo González BОценок пока нет
- Bloque IIIДокумент251 страницаBloque IIIGuillermo MazariegosОценок пока нет
- Núcleo Celular 2023Документ45 страницNúcleo Celular 2023José Javier Argüello castillo AОценок пока нет
- Semana 12 - Nucleo InterfasicoДокумент60 страницSemana 12 - Nucleo InterfasicoYENNIFER VILLAVICENCIO ROJASОценок пока нет
- NúcleoДокумент24 страницыNúcleoMafe Durán RangelОценок пока нет
- Clase 14 Nucleo y CromosomasДокумент53 страницыClase 14 Nucleo y CromosomasJose Carlos ALОценок пока нет
- Exposición Sobre El Nucleo de La Celula EucariotaДокумент14 страницExposición Sobre El Nucleo de La Celula Eucariotajohan laraОценок пока нет
- Capitulo 3 Nucleo Celular 1a Parte GeneralidadesДокумент90 страницCapitulo 3 Nucleo Celular 1a Parte GeneralidadesRonnie Reyes MartínezОценок пока нет
- Nucleo CelularДокумент23 страницыNucleo CelularMisael VargasОценок пока нет
- Exposision Nucleo 2Документ24 страницыExposision Nucleo 2JUAN ERNESTO ALVAREZ ROJASОценок пока нет
- 2 - Clase - Parte 1 - Conferencia Orientadora Núcleo y Ciclo Celular - CompressedДокумент32 страницы2 - Clase - Parte 1 - Conferencia Orientadora Núcleo y Ciclo Celular - CompressedNicole Aracelly PimboОценок пока нет
- 09 NucleoДокумент14 страниц09 NucleoYohanna BoteroОценок пока нет
- Diferencias Entre Procariota y EucariotaДокумент60 страницDiferencias Entre Procariota y EucariotaDaniel Eduardo Taborda (Médico veterinario)Оценок пока нет
- Núcleo Celular PDFДокумент38 страницNúcleo Celular PDFBety MartínОценок пока нет
- Núcleo Celular PDFДокумент37 страницNúcleo Celular PDFBety MartínОценок пока нет
- Clase 6 - NucleoДокумент23 страницыClase 6 - NucleoMontse Oria OrozcoОценок пока нет
- Nucleo y Nucleolo (INGRESO A MEDICINA UNC)Документ20 страницNucleo y Nucleolo (INGRESO A MEDICINA UNC)Barbi LindowОценок пока нет
- Nucleo y CromosomasДокумент43 страницыNucleo y CromosomasJEISSON DAVID CABOS SANCHEZОценок пока нет
- 01 Nucleo InterfasicoДокумент31 страница01 Nucleo InterfasicoGabo Yucra PatillaОценок пока нет
- Clase 6 - NucleoДокумент24 страницыClase 6 - NucleopinedobautistaedОценок пока нет
- Semana 7 - TeoriaДокумент23 страницыSemana 7 - TeoriajoselinОценок пока нет
- Núcleo CelularДокумент19 страницNúcleo CelularMaria Paula AranaОценок пока нет
- TEMA 5 NucleoloДокумент4 страницыTEMA 5 NucleoloALEX BERMUDOОценок пока нет
- TEMA 20 Nucleo y Nucleolo Mi GuiaДокумент10 страницTEMA 20 Nucleo y Nucleolo Mi Guiafrancisco hernandezОценок пока нет
- Semana 7 El Núcleo, Teoria 23-I PDFДокумент35 страницSemana 7 El Núcleo, Teoria 23-I PDFVALERYROSITA ATENCIOASCAMAОценок пока нет
- Monografia CB174 Amd 2016-Ii I GrupoДокумент67 страницMonografia CB174 Amd 2016-Ii I GrupoPercy Antony Huacarpuma HuayllaniОценок пока нет
- Clase 1. Constituyentes Del NucleoДокумент39 страницClase 1. Constituyentes Del NucleoDaniela CancinoОценок пока нет
- Tema - 11 Núcleo CelularДокумент57 страницTema - 11 Núcleo CelularErnesto Lluesma DropoОценок пока нет
- El Núcleo Celular 9Документ21 страницаEl Núcleo Celular 9MairaОценок пока нет
- Núcleo y NúcleoloДокумент10 страницNúcleo y NúcleoloXimena Huerta MonteroОценок пока нет
- Tema 19 Nucleo InterfasicoДокумент12 страницTema 19 Nucleo Interfasicochristina8reales8ca8Оценок пока нет
- Semana 7 Teoria - Nucleo CelularДокумент17 страницSemana 7 Teoria - Nucleo CelularEliza huamaniОценок пока нет
- 02-Cromatina-Est NuclearДокумент11 страниц02-Cromatina-Est NuclearSol NavarroОценок пока нет
- SUBTEMA 4 Trabajo BiologiaДокумент6 страницSUBTEMA 4 Trabajo BiologiaAmanda VeraОценок пока нет
- NÚCLEOДокумент14 страницNÚCLEOKaren Briggitte Bustamante VegaОценок пока нет
- NucleoДокумент28 страницNucleoRoberto ItzepОценок пока нет
- 13 NuclДокумент13 страниц13 NuclMarina BiagettiОценок пока нет
- Biología Semana 12 y 13Документ37 страницBiología Semana 12 y 13Fiorella SánchezОценок пока нет
- Documento Nucleo Usac 1 PDFДокумент29 страницDocumento Nucleo Usac 1 PDFRubí RecinosОценок пока нет
- Semana N °13: Núcleo-Estructura Y Función Cromosomas: Instituto de Educación Superior Tecnológico "Franklin RooseveltДокумент36 страницSemana N °13: Núcleo-Estructura Y Función Cromosomas: Instituto de Educación Superior Tecnológico "Franklin RooseveltMIKER ROCA TORRESОценок пока нет
- Sesión 07 - Biología CelularДокумент23 страницыSesión 07 - Biología CelularSILVINA JUDITH CABRERA MATTOSОценок пока нет
- Funcion Del NucleoloДокумент4 страницыFuncion Del NucleoloWalter Ruiz100% (1)
- 9-TEORÍA - Núcleo InterfásicoДокумент69 страниц9-TEORÍA - Núcleo InterfásicoJulian HuamaniОценок пока нет
- Estructura NuclearДокумент39 страницEstructura NuclearFernanda Veliz ArdilesОценок пока нет
- Núcleo y NucléoloДокумент12 страницNúcleo y NucléoloMarijose MezquiticОценок пока нет
- 3 NucleoДокумент25 страниц3 NucleoPaola Nathaly Nino100% (1)
- Wuolah Free App 1696328275979Документ5 страницWuolah Free App 1696328275979mangana.alba.miroОценок пока нет
- NUCLEOДокумент15 страницNUCLEOEsmi Esmi TexotlticitlОценок пока нет
- 8 - Nucleo CelularДокумент28 страниц8 - Nucleo CelularJuanCBОценок пока нет
- NUCLEOДокумент27 страницNUCLEOLUISINA BULACIOОценок пока нет
- El NucleoДокумент10 страницEl NucleoSmith Rels100% (1)
- DSC5831Документ76 страницDSC583120206394Оценок пока нет
- Nucleo, Ciclo Celular y Apoptosis. 2020Документ61 страницаNucleo, Ciclo Celular y Apoptosis. 2020otfelipe98Оценок пока нет
- Tema 9. El NúcleoДокумент12 страницTema 9. El NúcleoMarco MatellánОценок пока нет
- T-9 Nucleo Celular-Acidos Nucleicos-Prof. Anabel Gonzalez. OkДокумент28 страницT-9 Nucleo Celular-Acidos Nucleicos-Prof. Anabel Gonzalez. OkXimena Avila BerriosОценок пока нет
- Tríptico CofsafДокумент2 страницыTríptico CofsafCarlos ValdiviaОценок пока нет
- Expo MedicinaДокумент3 страницыExpo MedicinaCarlos ValdiviaОценок пока нет
- Patología-Enterocolitis InfecciosaДокумент37 страницPatología-Enterocolitis InfecciosaCarlos ValdiviaОценок пока нет
- Ejemplo de Historia DermatologíaДокумент5 страницEjemplo de Historia DermatologíaCarlos ValdiviaОценок пока нет
- Patología Del Aparato RespiratorioДокумент8 страницPatología Del Aparato RespiratorioJuan Cruz FenrirОценок пока нет
- Preguntasneumologia Casosclinicos 110320223035 Phpapp01Документ16 страницPreguntasneumologia Casosclinicos 110320223035 Phpapp01Brian Alex Herrera CheglioОценок пока нет
- SUEÑOДокумент2 страницыSUEÑOCarlos ValdiviaОценок пока нет
- Anexo IiДокумент16 страницAnexo IiCarlos ValdiviaОценок пока нет
- Estructura Articular Normal. 2015Документ25 страницEstructura Articular Normal. 2015Carlos ValdiviaОценок пока нет
- Fisiología HematoДокумент3 страницыFisiología HematoCarlos ValdiviaОценок пока нет
- Vocabulario (Algunas Palabras)Документ2 страницыVocabulario (Algunas Palabras)Carlos ValdiviaОценок пока нет
- Resumen Teorico2Документ1 страницаResumen Teorico2Carlos ValdiviaОценок пока нет
- Homosexual I DadДокумент20 страницHomosexual I DadCarlos ValdiviaОценок пока нет
- Seminario 5 Caso Clinico Obesidad 2014Документ2 страницыSeminario 5 Caso Clinico Obesidad 2014Carlos ValdiviaОценок пока нет
- VillaДокумент1 страницаVillaCarlos ValdiviaОценок пока нет
- Consentimiento InformadoДокумент1 страницаConsentimiento InformadoCarlos ValdiviaОценок пока нет
- EscalaCalidad Vida 2013Документ2 страницыEscalaCalidad Vida 2013Carlos ValdiviaОценок пока нет
- Adicciones Psicologicas Conceptualizacion e IntervencionДокумент7 страницAdicciones Psicologicas Conceptualizacion e IntervencionProspero PinoОценок пока нет
- Reporte Punto Focal Cusco-Parto VertiacalДокумент4 страницыReporte Punto Focal Cusco-Parto VertiacalCarlos ValdiviaОценок пока нет
- Solicit UdДокумент1 страницаSolicit UdCarlos ValdiviaОценок пока нет
- BioeticaДокумент20 страницBioeticaCarlos ValdiviaОценок пока нет
- ShockДокумент54 страницыShockCarlos ValdiviaОценок пока нет
- Etica 03Документ1 страницаEtica 03Carlos ValdiviaОценок пока нет
- Glandula Mamaria MedicinaДокумент72 страницыGlandula Mamaria MedicinaCarlos ValdiviaОценок пока нет
- Presentacion DM y Subsidios 2021Документ36 страницPresentacion DM y Subsidios 2021Luis Olivera ParedesОценок пока нет
- Práctica N°5 Proteinas IiiДокумент5 страницPráctica N°5 Proteinas IiiMarjorie Lizbeth P.Оценок пока нет
- FameДокумент2 страницыFameDafne LopezОценок пока нет
- Microbial Phylogeny and Diversity SSR RNA Sequence Analysis and BeyondДокумент12 страницMicrobial Phylogeny and Diversity SSR RNA Sequence Analysis and BeyondDeicy OrdoñezОценок пока нет
- Desarrollo Guia 1Документ11 страницDesarrollo Guia 1OHANA GAMERОценок пока нет
- Fnu U1 Atr DangДокумент4 страницыFnu U1 Atr Dangdaniel israel nava garciaОценок пока нет
- File 1599515788Документ13 страницFile 1599515788juan pablo escandón leguizamoОценок пока нет
- Drogas de Origen BiologicoДокумент17 страницDrogas de Origen Biologicosusy copa100% (1)
- Presentación 2 Introducción 2020Документ26 страницPresentación 2 Introducción 2020Luis AlvizuresОценок пока нет
- CT4to S1Документ4 страницыCT4to S1DIONISIO BAUTISTA GOMEZОценок пока нет
- La OvogénesisДокумент7 страницLa OvogénesisMaría JoséОценок пока нет
- Diapositivas de MicrobiologiaДокумент13 страницDiapositivas de MicrobiologiaVictor Samuel Marin OjedaОценок пока нет
- Grupo Mejoramiento Genético de Especies AutógamasДокумент1 страницаGrupo Mejoramiento Genético de Especies AutógamasJuan RiosОценок пока нет
- Programa Final 2019 BiotecnologíaДокумент112 страницPrograma Final 2019 BiotecnologíaRita AngelesОценок пока нет
- Prueba 1 CelulaДокумент4 страницыPrueba 1 CelulaCristina CostelaОценок пока нет
- Susana Cortés - Opciones en La Selección Espermática - II Simposio Reproducción Asistida QuirónДокумент43 страницыSusana Cortés - Opciones en La Selección Espermática - II Simposio Reproducción Asistida QuirónGrupo Hospitalario QuirónОценок пока нет
- Practica 2: Determinación in Vitro de La Permeación Gastroduodenal Del Salicilato de Sodio en RataДокумент15 страницPractica 2: Determinación in Vitro de La Permeación Gastroduodenal Del Salicilato de Sodio en RataGuillermo CasillasОценок пока нет
- Richard Escalona Semana 3Документ3 страницыRichard Escalona Semana 3Richard Escalona100% (1)
- Trabajo Semana 3 y 4Документ6 страницTrabajo Semana 3 y 4Oscar ReyesОценок пока нет
- El Gen Negro en El Pastor AlemánДокумент4 страницыEl Gen Negro en El Pastor AlemáncyberracingОценок пока нет
- 2.1.1. Membrana y Pared CelularДокумент27 страниц2.1.1. Membrana y Pared Celularjose adrian ojeda parraОценок пока нет
- L LLL N - NníwДокумент304 страницыL LLL N - NníwlauraОценок пока нет
- Reporte Introducción General A La Embriología Rocha Herrera Víctor ArturoДокумент2 страницыReporte Introducción General A La Embriología Rocha Herrera Víctor ArturoRocha ArturoОценок пока нет
- Bacterias 1Документ30 страницBacterias 1Ani Xiomara Armero PreciadoОценок пока нет
- Inhibidores de La ProteasaДокумент2 страницыInhibidores de La ProteasaClaudia PeñaОценок пока нет
- GediatriaДокумент6 страницGediatriaJuanito HernandezОценок пока нет
- Taller Organizacion Cuerpo Humano - 728578Документ6 страницTaller Organizacion Cuerpo Humano - 728578Vanessa OrtizОценок пока нет