Вам также может понравиться
- Заболевания щитовидной железы. Лечение и профилактика. 2-е изд.От EverandЗаболевания щитовидной железы. Лечение и профилактика. 2-е изд.Оценок пока нет
- EndoKrin 2013 01 062Документ8 страницEndoKrin 2013 01 062DariaОценок пока нет
- 120 / 80. Как привести давление в норму без таблеток (120 / 80. Kak privesti davlenie v normu bez tabletok)От Everand120 / 80. Как привести давление в норму без таблеток (120 / 80. Kak privesti davlenie v normu bez tabletok)Оценок пока нет
- Эндокринная система от @smart - medicДокумент17 страницЭндокринная система от @smart - medicalena.pashchenko17Оценок пока нет
- Patologia Endokrinnoy SistemyДокумент59 страницPatologia Endokrinnoy Sistemyvlad1nn999Оценок пока нет
- 43113Документ41 страница43113Мария ЛопатинаОценок пока нет
- ГИПОТАЛАМУС-ӨНЧИН ТАРХИНЫ БҮСИЙН ӨВЧНҮҮД PDFДокумент30 страницГИПОТАЛАМУС-ӨНЧИН ТАРХИНЫ БҮСИЙН ӨВЧНҮҮД PDFГанбат М.Оценок пока нет
- Rol de Distintas Sustancias en Los Procesos en El OrganismoДокумент40 страницRol de Distintas Sustancias en Los Procesos en El OrganismorosaОценок пока нет
- Gormonalnye Lekarstvennye SredstvaДокумент93 страницыGormonalnye Lekarstvennye SredstvaВалерия АлександровнаОценок пока нет
- стомат Лекции Частная гистологияДокумент55 страницстомат Лекции Частная гистологияАлиса СтолининаОценок пока нет
- АФО эндокр.систДокумент29 страницАФО эндокр.систМария ИвановаОценок пока нет
- Причины укорочения менструационного цикла - Женщинам и мужчинам рДокумент118 страницПричины укорочения менструационного цикла - Женщинам и мужчинам рAnna JuniedОценок пока нет
- Гинекологическая эндокринология, В.Н.ПрилепскаяДокумент264 страницыГинекологическая эндокринология, В.Н.ПрилепскаяОльга ШенягинаОценок пока нет
- Лекция 6 III курс. Фармакология гормональных препаратов. лечебный факультет.Документ14 страницЛекция 6 III курс. Фармакология гормональных препаратов. лечебный факультет.4everiamarmyОценок пока нет
- анатомияДокумент6 страницанатомияSulima TurdakunovaОценок пока нет
- Gumoralnaya_regulyatsiya_funktsiyДокумент44 страницыGumoralnaya_regulyatsiya_funktsiyryqhdqmqyqОценок пока нет
- Cпорт и эндокринная системаДокумент10 страницCпорт и эндокринная системаLidia ZamislaevОценок пока нет
- КЗ. ЖелезыДокумент6 страницКЗ. ЖелезыdesignerbeeniiОценок пока нет
- Voprosy 1-7Документ6 страницVoprosy 1-7ангелина кульчаковаОценок пока нет
- Metod Normal'Naya Fiziologiya Lechebnoe Delo-004Документ44 страницыMetod Normal'Naya Fiziologiya Lechebnoe Delo-004Анна РоманенкоОценок пока нет
- Lektsiya Dlya Studentov 5 Kursa Pediatricheskogo Fakulteta Ot 29.04.2020g. Nejroendokrinnye Sindromy. 1Документ61 страницаLektsiya Dlya Studentov 5 Kursa Pediatricheskogo Fakulteta Ot 29.04.2020g. Nejroendokrinnye Sindromy. 1Валерия СпивакОценок пока нет
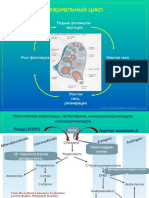
- Менструальный циклДокумент15 страницМенструальный циклАняОценок пока нет
- Biohimiya GormonovДокумент28 страницBiohimiya GormonovAndriy MisjuraОценок пока нет
- HistologieCurs (2) RuДокумент40 страницHistologieCurs (2) RuНаталья ОсоянуОценок пока нет
- ответы на экзамен с физы (теория)Документ115 страницответы на экзамен с физы (теория)kogerr86Оценок пока нет
- 93216Документ170 страниц93216tamarachitОценок пока нет
- Гормоны Белково-полипептидные (Копия)Документ15 страницГормоны Белково-полипептидные (Копия)shadiev.muxammadОценок пока нет
- гормоныДокумент4 страницыгормоныДана ОрумбаеваОценок пока нет
- 4 Endokrinologia 2017Документ11 страниц4 Endokrinologia 2017Enjoy vloggingОценок пока нет
- Шпаков АО Тиреоидная Система в Норме и При Сахарном Диабете 1-Го и 2-Го ТиповДокумент222 страницыШпаков АО Тиреоидная Система в Норме и При Сахарном Диабете 1-Го и 2-Го ТиповИван ЛебедевОценок пока нет
- Эндокринная система презентацияДокумент9 страницЭндокринная система презентацияGrigorosciuc CristinaОценок пока нет
- ПролактинДокумент6 страницПролактинSerj AlyОценок пока нет
- Первушина Е. В. - Железы и Гормоны. Эндокринная Система - (Семейная Энциклопедия Здоровья) - 2015Документ63 страницыПервушина Е. В. - Железы и Гормоны. Эндокринная Система - (Семейная Энциклопедия Здоровья) - 2015Новикова НатальяОценок пока нет
- Lektsia Fiziologia I Anatomia Organov ZHRSДокумент26 страницLektsia Fiziologia I Anatomia Organov ZHRSAnnaОценок пока нет
- Steroidy Razumny PodkhodДокумент60 страницSteroidy Razumny PodkhodIAMTNTОценок пока нет
- GormonyДокумент10 страницGormonyAnatolii IvankovОценок пока нет
- Starile de Hiperandrogenie in Practica Ginecologica Principii de Diagnostic Si TratamentДокумент8 страницStarile de Hiperandrogenie in Practica Ginecologica Principii de Diagnostic Si TratamentLndОценок пока нет
- Сро анатомияДокумент8 страницСро анатомияxnubirОценок пока нет
- Endokrinnye ZhelezyДокумент8 страницEndokrinnye ZhelezyxnubirОценок пока нет
- Zanyatie 17 Narushenie Polovogo Razvitia ГИНЕКОЛОГИЯДокумент13 страницZanyatie 17 Narushenie Polovogo Razvitia ГИНЕКОЛОГИЯLester O'connorОценок пока нет
- LektsiiДокумент114 страницLektsiiGheorghe SecuОценок пока нет
- Анатомическое строение гипоталамусаДокумент10 страницАнатомическое строение гипоталамусаЭрнис ЭралиевОценок пока нет
- эндокринная система_ПетлюкДокумент3 страницыэндокринная система_ПетлюкЕкатерина ПетлюкОценок пока нет
- Возраст инволюцийДокумент14 страницВозраст инволюцийІрина БазікалоОценок пока нет
- Гипопитуитаризм (презентация)Документ33 страницыГипопитуитаризм (презентация)lupinotuum63Оценок пока нет
- Endokrinologia BalabolkinДокумент503 страницыEndokrinologia BalabolkinIrina SumeicoОценок пока нет
- 148 673Документ24 страницы148 673aОценок пока нет
- Сухарев-Психология интеллектаДокумент138 страницСухарев-Психология интеллектаYaroslav SolovkoОценок пока нет
- 35. Система питанияДокумент7 страниц35. Система питанияВалерия АлександровнаОценок пока нет
- Био ОГЭ - 31, Эндокринная система PDFДокумент45 страницБио ОГЭ - 31, Эндокринная система PDFДаниил ПалиховОценок пока нет
- 709633Документ37 страниц709633jaroslav.mudryjОценок пока нет
- 2012 Гормональные Препараты - КопияДокумент35 страниц2012 Гормональные Препараты - КопияНикита АлександровичОценок пока нет
- Cerebrolizin Geroprot 2020Документ8 страницCerebrolizin Geroprot 2020I TОценок пока нет
- Saatov 6 13Документ12 страницSaatov 6 13Lucia BogОценок пока нет
- Патофизиология критических состоянийДокумент423 страницыПатофизиология критических состоянийDiana IvanovaОценок пока нет
- РефератДокумент14 страницРефератTeresa TumanyanОценок пока нет
- Половые гормоны и репродуктивноеДокумент8 страницПоловые гормоны и репродуктивноеАнтон ПосныйОценок пока нет
- 2 - Система Регуляций ФункцийДокумент36 страниц2 - Система Регуляций ФункцийOushminalda SmithОценок пока нет
- Регуляция процессов метаболизма у бактерийДокумент135 страницРегуляция процессов метаболизма у бактерийЮлияОценок пока нет
- Эндокринная гинекологияДокумент301 страницаЭндокринная гинекологияМария ВасильеваОценок пока нет
- The Mama Natural Week-by-Week Guide to Pregnancy and ChildbirthОт EverandThe Mama Natural Week-by-Week Guide to Pregnancy and ChildbirthРейтинг: 4.5 из 5 звезд4.5/5 (21)
- Wordslut: A Feminist Guide to Taking Back the English LanguageОт EverandWordslut: A Feminist Guide to Taking Back the English LanguageРейтинг: 4.5 из 5 звезд4.5/5 (82)
- People to Be Loved: Why Homosexuality Is Not Just an IssueОт EverandPeople to Be Loved: Why Homosexuality Is Not Just an IssueРейтинг: 4.5 из 5 звезд4.5/5 (23)
- Period Power: Harness Your Hormones and Get Your Cycle Working For YouОт EverandPeriod Power: Harness Your Hormones and Get Your Cycle Working For YouРейтинг: 4 из 5 звезд4/5 (25)
- Anatomy 101: From Muscles and Bones to Organs and Systems, Your Guide to How the Human Body WorksОт EverandAnatomy 101: From Muscles and Bones to Organs and Systems, Your Guide to How the Human Body WorksРейтинг: 4.5 из 5 звезд4.5/5 (19)
- Unwanted: How Sexual Brokenness Reveals Our Way to HealingОт EverandUnwanted: How Sexual Brokenness Reveals Our Way to HealingРейтинг: 4.5 из 5 звезд4.5/5 (30)
- Sexual Ethics and Islam: Feminist Reflections on Qur'an, Hadith, and JurisprudenceОт EverandSexual Ethics and Islam: Feminist Reflections on Qur'an, Hadith, and JurisprudenceРейтинг: 4.5 из 5 звезд4.5/5 (8)
- The Only Pregnancy Book You'll Ever Need: An Expectant Mom's Guide to EverythingОт EverandThe Only Pregnancy Book You'll Ever Need: An Expectant Mom's Guide to EverythingРейтинг: 4.5 из 5 звезд4.5/5 (3)
- Dude, You're a Fag: Masculinity and Sexuality in High School, With a New PrefaceОт EverandDude, You're a Fag: Masculinity and Sexuality in High School, With a New PrefaceРейтинг: 3.5 из 5 звезд3.5/5 (33)
- Gay Girl, Good God: The Story of Who I Was, and Who God Has Always BeenОт EverandGay Girl, Good God: The Story of Who I Was, and Who God Has Always BeenРейтинг: 4.5 из 5 звезд4.5/5 (139)
- Your Baby, Your Way: Taking Charge of your Pregnancy, Childbirth, and Parenting Decisions for a Happier, Healthier FamilyОт EverandYour Baby, Your Way: Taking Charge of your Pregnancy, Childbirth, and Parenting Decisions for a Happier, Healthier FamilyРейтинг: 4 из 5 звезд4/5 (8)
- DIY Porn Handbook: A How-To Guide to Documenting Our Own Sexual RevolutionОт EverandDIY Porn Handbook: A How-To Guide to Documenting Our Own Sexual RevolutionРейтинг: 1 из 5 звезд1/5 (1)
- What to Expect for First-Time Moms: The Ultimate Beginners Guide While Expecting, Everything You Need to Know for a Healthy Pregnancy, Labor, Childbirth, and Newborn - A New Mom’s Survival HandbookОт EverandWhat to Expect for First-Time Moms: The Ultimate Beginners Guide While Expecting, Everything You Need to Know for a Healthy Pregnancy, Labor, Childbirth, and Newborn - A New Mom’s Survival HandbookРейтинг: 4.5 из 5 звезд4.5/5 (8)
- Habits of a Happy Brain: Retrain Your Brain to Boost Your Serotonin, Dopamine, Oxytocin, & Endorphin LevelsОт EverandHabits of a Happy Brain: Retrain Your Brain to Boost Your Serotonin, Dopamine, Oxytocin, & Endorphin LevelsРейтинг: 4.5 из 5 звезд4.5/5 (151)
- The Hormone Reset Diet: Heal Your Metabolism to Lose Up to 15 Pounds in 21 DaysОт EverandThe Hormone Reset Diet: Heal Your Metabolism to Lose Up to 15 Pounds in 21 DaysРейтинг: 4 из 5 звезд4/5 (40)
- What's Happening to My Body? Book for Girls: Revised EditionОт EverandWhat's Happening to My Body? Book for Girls: Revised EditionРейтинг: 4 из 5 звезд4/5 (7)
- Healing the Thyroid with Ayurveda: Natural Treatments for Hashimoto's, Hypothyroidism, and HyperthyroidismОт EverandHealing the Thyroid with Ayurveda: Natural Treatments for Hashimoto's, Hypothyroidism, and HyperthyroidismРейтинг: 4.5 из 5 звезд4.5/5 (8)
- $pread: The Best of the Magazine that Illuminated the Sex Industry and Started a Media RevolutionОт Everand$pread: The Best of the Magazine that Illuminated the Sex Industry and Started a Media RevolutionRachel AimeeРейтинг: 4.5 из 5 звезд4.5/5 (5)
- The Fifth Vital Sign: Master Your Cycles & Optimize Your FertilityОт EverandThe Fifth Vital Sign: Master Your Cycles & Optimize Your FertilityРейтинг: 4.5 из 5 звезд4.5/5 (12)
- The Vagina Bible: The Vulva and the Vagina: Separating the Myth from the MedicineОт EverandThe Vagina Bible: The Vulva and the Vagina: Separating the Myth from the MedicineРейтинг: 4.5 из 5 звезд4.5/5 (27)