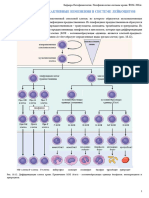
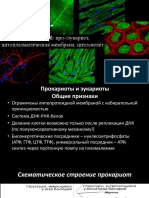
Вам также может понравиться
- Система гуморального иммунитетаДокумент11 страницСистема гуморального иммунитетаMazurencoIanaОценок пока нет
- 59663Документ13 страниц59663dietox.ruОценок пока нет
- презент 1Документ32 страницыпрезент 1lovehappyОценок пока нет
- VirusyДокумент48 страницVirusyiamannett19Оценок пока нет
- ИММУНОЛОГИЯДокумент19 страницИММУНОЛОГИЯAhmed AymanОценок пока нет
- ЛАТ - л4-ифа1Документ25 страницЛАТ - л4-ифа1Дима ВласюкОценок пока нет
- ImmunologiayayayaДокумент48 страницImmunologiayayayaahmedelghallabyОценок пока нет
- план и вопросы к занятиям леч фак 2020Документ14 страницплан и вопросы к занятиям леч фак 2020elsaman00Оценок пока нет
- 14 Иммун 5 РеакцииДокумент19 страниц14 Иммун 5 РеакцииDjgernaut DotinОценок пока нет
- Лекция №1Документ54 страницыЛекция №1Alexandra BaluhatayaОценок пока нет
- Адельман Д. - Клиническая иммунология и аллергологияДокумент220 страницАдельман Д. - Клиническая иммунология и аллергологияIrina Moldovan100% (1)
- Biologicheski MembranyДокумент8 страницBiologicheski MembranyNoir BlackОценок пока нет
- 11 ИммДокумент35 страниц11 ИммDjgernaut DotinОценок пока нет
- Mikra DocxДокумент75 страницMikra DocxДарья Константинова100% (1)
- Gistologia Ekzamen2 1Документ48 страницGistologia Ekzamen2 1Александр ФеоктистовОценок пока нет
- Гистология супортДокумент190 страницГистология супортНаталья ОсоянуОценок пока нет
- Лекция Иммунная система и её функцииДокумент35 страницЛекция Иммунная система и её функцииАмир СадретдиновОценок пока нет
- 123906Документ26 страниц123906dietox.ruОценок пока нет
- BioДокумент109 страницBioxtdksnm825Оценок пока нет
- Компартментизация клеток эукариотДокумент18 страницКомпартментизация клеток эукариотЮлияОценок пока нет
- 2-6 ВВИГДокумент9 страниц2-6 ВВИГOlgaОценок пока нет
- 1 Kollok - Preparaty-1Документ21 страница1 Kollok - Preparaty-164fknjvqkvОценок пока нет
- Teorii Immuniteta. Immunologicheskaya Pamyat. Immunologicheskaya Tolerantnost. Regulyatsiya Immuniteta.Документ17 страницTeorii Immuniteta. Immunologicheskaya Pamyat. Immunologicheskaya Tolerantnost. Regulyatsiya Immuniteta.bagdauletamangeldi1Оценок пока нет
- BiologiaДокумент73 страницыBiologiaBanda BandovОценок пока нет
- ИммунологияДокумент56 страницИммунологияisaacoff013Оценок пока нет
- Immune GM Ru 2022Документ27 страницImmune GM Ru 2022kabulkabulovich5Оценок пока нет
- PresentatsiyaДокумент22 страницыPresentatsiyaShakhbozОценок пока нет
- WBC GM Ru 2024Документ14 страницWBC GM Ru 2024kabulkabulovich5Оценок пока нет
- GormonyДокумент10 страницGormonyAnatolii IvankovОценок пока нет
- иммунитет 2Документ70 странициммунитет 2dinakuchukovaОценок пока нет
- IMMUNOLOGIYa Lektsia 2 Immunnaya Sistema DocДокумент16 страницIMMUNOLOGIYa Lektsia 2 Immunnaya Sistema Docarthurbogning0Оценок пока нет
- 2-практическое занятиеДокумент15 страниц2-практическое занятиеlazikotoshpulatovaОценок пока нет
- PHP Z3 Oco XДокумент14 страницPHP Z3 Oco Xsarvinoz51Оценок пока нет
- лабораторная работа 5Документ4 страницылабораторная работа 5Арнаут МихаелаОценок пока нет
- Lekcziya 3 Morfologiya I Strukturno Funkczionalnaya OrganizacziyaДокумент54 страницыLekcziya 3 Morfologiya I Strukturno Funkczionalnaya OrganizacziyaЕкатерина НовиковаОценок пока нет
- Immunologia 2020 - PostnikovaДокумент257 страницImmunologia 2020 - PostnikovaNikitaОценок пока нет
- анатомо-физиологические особенности иммунной системыДокумент54 страницыанатомо-физиологические особенности иммунной системыRomanОценок пока нет
- Биология клеткиДокумент13 страницБиология клеткиBanda BandovОценок пока нет
- трансляция как завершающий этап биосинтезаДокумент9 страництрансляция как завершающий этап биосинтезаРахым ХасеновОценок пока нет
- Лекция 1Документ24 страницыЛекция 1Համլետ ԿարայանОценок пока нет
- имункаДокумент11 страницимункаanna.obertun2001Оценок пока нет
- органоидыДокумент25 страницорганоидыaisha zaripovaОценок пока нет
- Презентация " Органоиды клетки"Документ26 страницПрезентация " Органоиды клетки"zamirov.ilyazОценок пока нет
- иммунитет 1 лектор МишинаДокумент18 странициммунитет 1 лектор МишинаAuthentic IdiotОценок пока нет
- Л 6.Обмен Веществ и Энергии в Клетке (2) -1Документ41 страницаЛ 6.Обмен Веществ и Энергии в Клетке (2) -1xevildoersОценок пока нет
- Реферат КЛДДокумент14 страницРеферат КЛДNataliyaОценок пока нет
- Fiza - 4 - 1 - Modul 2Документ38 страницFiza - 4 - 1 - Modul 2Melanie Quiroga DelgadoОценок пока нет
- БиомембраныДокумент12 страницБиомембраныДжама МехтиеваОценок пока нет
- ДокументДокумент9 страницДокументirinka051812Оценок пока нет
- фагоцитоз микра (копія)Документ5 страницфагоцитоз микра (копія)daria.mrzv20Оценок пока нет
- Лабораторная работа 17Документ3 страницыЛабораторная работа 17Арнаут МихаелаОценок пока нет
- Молекулярные механизмы процессов энергетического сопряженияДокумент12 страницМолекулярные механизмы процессов энергетического сопряженияГерман НикифоровОценок пока нет
- 10строение Клетки - КопияДокумент24 страницы10строение Клетки - КопияajdajbajmamatovaОценок пока нет
- 40497Документ32 страницы40497dietox.ruОценок пока нет
- Презентация Microsoft PowerPointДокумент8 страницПрезентация Microsoft PowerPointАнастасия НегодаОценок пока нет
- Одномембранные орг11Документ4 страницыОдномембранные орг11Роман Кий (Dragon)Оценок пока нет
- ИммунологияДокумент52 страницыИммунологияsevdaismailova13Оценок пока нет
- 3. Биологические Мембраны-2020Документ60 страниц3. Биологические Мембраны-2020Ruslan KinevОценок пока нет
- Углевод-белковые комплексы.Биохимия межклеточного матрикса.Документ30 страницУглевод-белковые комплексы.Биохимия межклеточного матрикса.nazarovmuhammad1131Оценок пока нет
- Невидимый страж: Как иммунитет защищает нас от внешних и внутренних угрозОт EverandНевидимый страж: Как иммунитет защищает нас от внешних и внутренних угрозОценок пока нет
- ImmulightДокумент5 страницImmulightFuadОценок пока нет
- Immunology SynapseДокумент105 страницImmunology SynapseDmitry100% (1)
- цмвиДокумент72 страницыцмвиxiadneyaa xiadneyaaОценок пока нет
- Preturi Laborator RUДокумент11 страницPreturi Laborator RUVulpe EugeniaОценок пока нет
- Mikra DocxДокумент75 страницMikra DocxДарья Константинова100% (1)
- ImmunolekДокумент14 страницImmunolekp28dn9ppdcОценок пока нет